 Экологические основы интегрированной защиты растений: учебник
Экологические основы интегрированной защиты растений: учебник

 ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ

 ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ


3.4.1. Источники воспроизводства вредных организмов в агроэкосистемах (тактика Р)
Способность размножаться, или производить новое поколение особей любого вида, является главной фундаментальной функцией живых организмов. В процессе размножения происходит непрерывная передача информации от родителей к потомкам, что позволяет воспроизводить признаки жизненного цикла вида. Вид может существовать только в том случае, если каждое новое поколение будет производить больше потомков, чем было родительских особей, принимавших участие в их создании.
Каждый организм (в форме видовой популяции) при размножении получает набор генетических программ, которые в разной степени реализуются в процессе индивидуального развития (онтогенеза), проявляясь затем в форме фенотипических признаков.
Существует два основных типа размножения – бесполое и половое, признаки которых приведены в табл. 26.
Бесполое размножение фитопатогенов (возбудители ржавчинных заболеваний, септориоза), фитофагов (тли, клещи, белокрылки) в результате митоза может привести к массовой численности вредных организмов и значительным потерям урожая сельскохозяйственных культур и снижению качества продукции.
Под митозом понимают такое деление клеточного ядра, при котором образуются два дочерних ядра с набором хромосом, идентичных набору родительской клетки. Самое важное событие во время митоза – равное распределение между двумя дочерними клетками удвоившихся хромосом. |
Таблица 26
Сравнение признаков бесполого и полового размножения
Бесполое размножение |
Половое размножение |
Одна родительская особь Гаметы не образуются |
Обычно две родительские особи Образуются гаплоидные гаметы, ядра которых сливаются, происходит оплодотворение и образование диплоидных зигот |
Мейоз отсутствует, присутствует митоз |
На какой-то стадии жизненного цикла происходит мейоз, что препятствует удвоению хромосом в каждом поколении |
Потомки идентичны родительским особям |
Потомки не идентичны родительским особям. У них наблюдается генетическая изменчивость в результате генетической рекомбинации |
Характерно для растений, низших животных, микроорганизмов |
Характерно для большинства растений и животных |
Часто приводит к быстрому созданию большого числа потомков |
Менее быстрое увеличение численности |
У растений митоз происходит главным образом в меристемах, у животных – в различных тканях и участках организма. Значение митоза состоит в обеспечении генетической стабильности организма. Учитывая механизмы репликации ДНК при митозе, возможность генетической изменчивости у организмов с бесполым размножением (например, у грибов – основных возбудителей болезней растений) очень мала. Любая видимая изменчивость при бесполом размножении организмов чаще обусловлена мутациями и воздействием факторов внешней среды.
Генетическая идентичность потомков при бесполом размножении вредных организмов (бактерий, грибов, ряда насекомых) гарантирует их совместимость с генотипом растений-хозяев. Это позволяет видовым популяциям в короткие сроки увеличить в значительной мере численность популяций. Так, в течение 2-х месяцев популяция возбудителя фитофтороза картофеля в результате продуцирования зооспор и конидий при благоприятных гидротермических условиях возрастает в 5·1017 раз на 1 га в сутки, а популяция урединиоспор возбудителя бурой ржавчины пшеницы – в 3,2·103 раз на 1 га в сутки. Это приводит к вспышке ЭП, а при наличии больших площадей генетически совместимых восприимчивых сортов картофеля и пшеницы, а также благоприятных гидротермических условиях для воздушно-капельного механизма передачи фитопатогенов – к эпифитотии и панфитотии. Поэтому К.М. Степанов отмечал, что «проблема грибных эпифитотий является, в сущности, проблемой массового размножения возбудителя».
Генетическая изменчивость при бесполом размножении, которая приводит к формированию новых рас и биотипов, преодолевающих устойчивость сортов к возбудителям болезней, обеспечивается в основном мутациями, частота которых может возрастать при резком изменении условий окружающей среды, например, при применении фунгицидов. В настоящее время возникновение мутаций известно у представителей всех классов фитопатогенных грибов, например у Ustilago maydis (базидиальные грибы), Bipolaris sorokiniema (несовершенные).
В отличие от бесполого, при половом размножении образуются гаметы зрелых женских и мужских половых клеток, содержащих в результате мейоза гаплоидный набор хромосом. Во время мейоза в результате случайного расхождения хромосом по гаметам, а также обмена гомологичными частями хромосом (кроссинговера) возникают новые комбинации генов.
Слияние содержащихся в гаметах гаплоидных ядер называют оплодотворением, которое приводит к образованию диплоидной зиготы с двойным набором хромосом. Таким образом, при половом размножении видовых популяций происходит чередование диплоидных и гаплоидных фаз, обусловливая непрерывность жизненного цикла и лучшую выживаемость особей в популяции.
Общая плодовитость таксономических групп вредных организмов, размножающихся половым способом (насекомые, грызуны, сорные растения), значительно ниже, чем у бактерий и грибов, размножающихся преимущественно бесполым способом. Так, средняя плодовитость большинства насекомых составляет примерно 200 яиц, в то время как грибов – 2·105. При этом половое размножение происходит, как правило, один раз на протяжении жизненного цикла, в то время как бесполое – многократно.
Половое размножение отличается большей изменчивостью, обусловливая формирование генетически разнообразных потомков вредных организмов менее совместимых с сортами сельскохозяйственных культур. Тем самым бесполое размножение выполняет функцию увеличения плотности преимущественного генетически идентичных популяций, а половое – гетерогенных.
Вредные организмы приурочены (кроме грызунов) к размножению в популяции растений-хозяев, заняв различные экологические ниши (табл. 27).
Таблица 27
Экологические ниши размножения вредных организмов в агро- и естественных экосистемах
Таксономические группы вредных организмов |
Организм восприимчивых растений |
Вне организма, в популяции восприимчивых растений |
За пределами популяции восприимчивых растений |
|||
клетки |
ткани органов |
сосудисто-проводящая система |
||||
вегетативных |
генеративных |
|||||
Вирусы |
+ |
+ |
+ |
+ |
- |
+ |
Бактерии |
+ |
+ |
+ |
+ |
- |
- |
Грибы |
+ |
+ |
+ |
+ |
- |
- |
Нематоды |
- |
+ |
+ |
- |
- |
- |
Насекомые |
- |
+ |
+ |
- |
+ |
- |
Грызуны |
- |
- |
- |
- |
- |
+ |
Сорные растения: |
||||||
свободноживущие |
- |
- |
- |
- |
+ |
- |
паразитические |
- |
+ |
- |
- |
- |
- |
Фитопатогены (вирусы, бактерии, грибы) размножаются в клетках и тканях вегетативных и генеративных органов, а также в сосудисто-проводящей системе восприимчивых растений в агро- и естественных экосистемах; насекомые – на (реже в) тканях вегетативных и генеративных органов, а также около растений, размещая яйцекладку на поверхности или в верхнем слое почвы. Свободноживущие сорные растения размножаются преимущественно в посевах культурных растений (агроценозах), а также вне агроэкосистем, заселяя пустыри, селитебные и хозяйственные территории, транспортные магистрали, линии электропередач и др. Паразитические сорняки размножаются на органах растений, присасываясь к их поверхности. Грызуны, хотя и размножаются в норах на полях озимых зерновых культур и многолетних трав, но предпочитают целинные и залежные участки (естественные экосистемы). Вирусы, кроме восприимчивых растений-хозяев могут размножаться в организме насекомых, а возможно, нематод и грибов. Итак, для подавляющего большинства вредных организмов популяции восприимчивых культурных растений являются источников воспроизводства (размножения) их популяций. Специалист по защите растений должен иметь четкое представление о местах массового размножения вредных организмов в агро- и естественных экосистемах, а, следовательно, первичных эпифитотических очагах. Поскольку эпифитотии и массовое размножение начинается со стартовых площадок эпифитотических очагов, возникает необходимость в их первичной локализации. В ином случае отсутствие мониторинга мест размножения вредных организмов в агро- и естественных экосистемах тормозит и делает невозможным развитие экологического направления защиты растений в плане ограниченного применения пестицидов. Поэтому на данном этапе развития науки, кроме традиционного мониторинга численности вредных организмов возникает необходимость мониторинга объектов их массового размножения в агро- и естественных экосистемах. Это вызывает в свою очередь необходимость научного определения источника воспроизводства (источника возбудителя болезней) вредных организмов.
Источник воспроизводства вредных организмов (источник ВВО, или источник возбудителя болезни) – это объект, который служит местом их естественного, выработанного в процессе эволюции, размножения, из которого они выделяются (попадают) жизнеспособными в окружающие стации. Совокупность источников ВВО на данной территории составляет их резервуар. |
Анализ жизненного цикла фитопатогенов и фитофагов показал, что они различаются по количеству и качеству источников воспроизводства в агроэкосистемах (рис. 14).
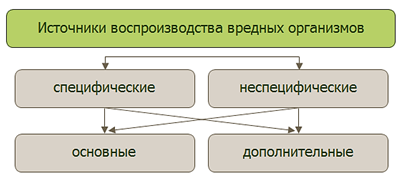
Рис. 14. Классификация источников воспроизводства вредных организмов
Источниками воспроизводства могут быть сельскохозяйственные культуры, сорные растения, представители дикой флоры. Их роль в увеличении численности вредных организмов различна. По этому признаку их классифицируют на четыре группы: специфические, неспецифические, основные, дополнительные.
К специфическим отнесены такие источники ВВО, которые выработаны в процессе эволюции и обеспечивают постоянное воспроизводство вредных организмов в агро- и естественных экосистемах (в природе). Так, постоянное воспроизводство возбудителей бурой ржавчины, корневых гнилей зерновых культур происходит на дикорастущих многолетних злаках – пырее ползучем, тимофеевке, костреце и др. Специфические источники служат стартовой площадкой для формирования природных эпифитотических очагов, особенно наземно-воздушных вредных организмов (возбудители бурой и стеблевой ржавчины, саранчовые, луговой мотылек и др.).
Неспецифические источники воспроизводства вредных организмов являются случайными, связанными часто с хозяйственной деятельностью человека. Например, известно, что в процессе механизированной уборки кукурузы на силос теряется много ее фитомассы. Она осыпается на почву, представляя собою благоприятную питательную среду для почвенных фитопатогенов, численность которых возрастает во много раз. Это объясняет случаи высокой заселенности почв возбудителем корневых гнилей Bipolaris sorokiniana после такого, по сути, фитосанитарного предшественника, как кукуруза.
К основным источникам воспроизводства относятся такие, которые служат местом массового размножения вредных организмов в агро- или (реже) естественных экосистемах. Именно основные источники ВВО обусловливают массовую численность вредных организмов в агроэкосистемах и эпифитотические вспышки болезней растений или эпифитотии.
Основные источники ВВО находятся, как правило, в агроэкосистемах. Ими служат восприимчивые сорта и культуры. По нашим данным, 61 % основных источников размножения возбудителей широко распространенных и вредоносных болезней зерновых культур (корневые гнили, септориоз, головневые заболевания, спорынья), зернобобовых культур (корневые гнили, аскохитоз), картофеля (ризоктониоз, фитофтороз, альтернариоз) и др. находятся в агроэкосистемах, а 39 % источников размножения (ржавчинные заболевания) на (в) – в многолетних злаках целинных и залежных участках естественных экосистем.
|
Для регулирования численности вредных организмов и проявления ЭП в агроэкосистемах решающее значение имеет выяснение основных источников, где происходит массовое размножение популяций вредных организмов. |

Например, массовое размножение возбудителя гельминтоспориозной корневой гнили происходит на ячмене. Тем самым происходит массовое заселение фитопатогеном почвы, предназначенной под яровую пшеницу – главную продовольственную культуру. Вследствие этого возникает необходимость фитопатологического мониторинга агроценозов после уборки ячменя с целью принятия решения о целесообразности возделывания яровой пшеницы в конкретном севообороте. Особенно это важно при возделывании твердой яровой пшеницы, зерно которой при сильном поражении чернотой зародыша непригодно для производства макарон и манной крупы.
|
Таким образом, все вредные организмы имеют в агроэкосистемах объекты массового размножения – восприимчивые культуры, сорта, сорные растения. Их выявление и локализация которых одна из первостепенных задач систем ИЗР. Вторая задача – выявление и нейтрализация факторов передачи (сохранения, расселения) вредных организмов – тактики В. |



