 Экологические основы интегрированной защиты растений: учебник
Экологические основы интегрированной защиты растений: учебник

 ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ

 ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ


3.1. Формирование сообществ вредных организмов в агроэкосистемах
Под методологией понимают учение о принципах построения, формах и способах научно-познавательной деятельности. |
Для разработки систем ИЗР необходимо знать следующие научные положения:
 закономерности формирования сообществ и экологических ниш вредных организмов в агроэкосистемах;
закономерности формирования сообществ и экологических ниш вредных организмов в агроэкосистемах;
 взаимодействия вредных организмов с сельскохозяйственными растениями, друг с другом и с другими компонентами агроэкосистем, особенно энтомофагами, антагонистами;
взаимодействия вредных организмов с сельскохозяйственными растениями, друг с другом и с другими компонентами агроэкосистем, особенно энтомофагами, антагонистами;
 функциональную модель эпифитотического процесса и методологию рассмотрения действия и взаимодействия факторов, влияющих на численность вредных организмов и развитие болезней;
функциональную модель эпифитотического процесса и методологию рассмотрения действия и взаимодействия факторов, влияющих на численность вредных организмов и развитие болезней;
 экологическую классификацию вредных организмов, как методологическую основу для создания условий в агроэкосистемах благоприятных для растений и неблагоприятных – для вредных организмов.
экологическую классификацию вредных организмов, как методологическую основу для создания условий в агроэкосистемах благоприятных для растений и неблагоприятных – для вредных организмов.
Термин «экосистема» введен в науку английским экологом А. Тэнсли в 1935 г. С тех пор это понятие превратилось в основную функциональную единицу экологии.
|
Под экосистемой понимают любое сообщество живых организмов и неживых компонентов (органических, неорганических), составляющих функциональное целое, в котором может осуществляться круговорот веществ. |

С эволюционно-экологической точки зрения различают два типа экосистем: естественные и агроэкосистемы.
До возникновения земледелия все экосистемы были естественными, или природными. Они возникли в процессе сопряженной эволюции биологических сообществ с природными абиотическими факторами окружающей среды и представлены в настоящее время целинными и залежными участками, а также лесами, колками, болотами, поймами рек и другими участками земной поверхности, которые функционируют в природе без существенного вмешательства человека. Естественные экосистемы отличаются сбалансированностью, подвижным равновесием (гомеостаз) и функциональной саморегуляцией их компонентов. Они прошли длительный путь сукцессионных изменений.
Под сукцессией понимают упорядоченный процесс смены одних биологических сообществ другими. |
Сукцессионные процессы называются первичными, если они возникают на новых, лишенных жизни местообитаниях и вторичными – при возникновении на нарушенных местообитаниях. Конечным пунктом сукцессий является формирование климаксовых (уравновешенных, стабильных) сообществ. С этой точки зрения в агроэкосистемах происходят вторичные сукцессии, которые возникли и вновь возникают по мере развития земледелия.
В настоящее время в мировом сельском хозяйстве используется около 4,7 млрд. га сельскохозяйственных угодий, в том числе 1,4 млрд. га пашни, которые можно рассматривать как агроэкосистемы различных типов, подверженных на 76 % территорий температурному, водному, минеральному или фитосанитарному стрессу.
Во всех случаях под термином "агроэкосистема" понимают совокупность растений, животных и микроорганизмов, их местообитания, в разной степени измененные и используемые в процессе трудовой деятельности человека. |
В сельскохозяйственной литературе термин «агроэкосистема» рассматривается как синоним терминов «агробиогеоценоз», «агроценоз». Существо вопроса сводится к анализу взаимодействия биологических сообществ и связанных с ними абиотических факторов чаще всего в пределах такого сельскохозяйственного объекта как поле (посев сельскохозяйственной культуры или ее агроценоз), севооборота, ландшафта. Эти объекты являются разными территориальными вариантами агроэкосистем, а поэтому тесно взаимосвязаны друг с другом. В их изучении и регулировании много общего, хотя есть и отличия.
В защите растений агроэкосистемы лучше изучены на уровне посевов сельскохозяйственных культур и, частично – севооборотов. Это обусловлено господствующими в течение прошлого столетия системами земледелия – травопольной, паропропашной, почвозащитной, интенсивной, которые были ориентированы преимущественно на преобразование пашни. При переходе к адаптивно-ландшафтной системе земледелия возникает необходимость рассмотрения взаимодействия биологических сообществ, включая вредные организмы, шире – на уровне агроландшафтов, которые характеризуются различным рельефом, высотой над уровнем моря, сетью рек и ручьев. Компоненты ландшафта как экосистемы находятся в системной связи друг с другом, выступая как единое целое с характерным гидротермическим режимом, почвенно-климатическими условиями, орографией (гр. oros – гора, grapho – пишу) сообществом растений, животных и микроорганизмов, сформировавшихся в процессе эволюции (естественные ландшафты) и измененных по мере развития земледелия (агроландшафты).
За всю историю земледелия не отмечено появления новых видов вредных организмов в агроэкосистемах: все они были известны как компоненты естественных экосистем. Антропогенное влияние затронуло преимущественно процессы формо- и расообразования, адаптации тактик жизненного цикла, повлияло на соотношение и взаимоотношения видовых популяций в сообществах агроэкосистем.
В современных агроэкосистемах насчитывают примерно 8500 возбудителей болезней, около 10 тыс. видов фитофагов, 1500 нематод и более 2000 видов сорняков, наносящих экономически значимый ущерб сельскохозяйственным культурам. Число потенциально опасных вредных организмов в агроэкосистемах превышает 100 тыс. видов.
Формирование сообществ вредных организмов в агроэкосистемах происходило и происходит в настоящее время двумя путями: миграцией их из центров происхождения культурных растений (первичных, вторичных), в процессе развития торговли и обмена и переселением из естественных аборигенных (местных) экосистем.
До возникновения земледелия все виды вредных организмов входили в состав биологических сообществ естественных экосистем. При этом первичные ареалы многих специализированных видов совпадали с центрами происхождения культурных растений (табл. 18).
Таблица 18
Центры происхождения культурных растений
Ареал |
Происхождение культуры |
|
первичное |
вторичное |
|
Юго-западная Азия от Кавказа и востока Малой Азии до Памира |
Пшеница, рожь, овес, двухрядный ячмень, мелкосеменная форма льна, чечевица, бобы, горох, люцерна, сурепица, лук репчатый, яблоня, груша, слива, вишня, черешня |
|
Горные ландшафты и побережье средиземноморских стран |
Люпин, клевер, сахарная и кормовая свекла, капуста, рапс, морковь |
Горох, бобы, лен, чечевица, петрушка, сельдерей |
Абиссинское нагорье |
Пшеница, чечевица, горох, ячмень, кофе |
|
Африка |
Сорго |
|
Юго-Восточная Азия Индия |
Ячмень, персик, абрикос |
|
Мексика, Гватемала |
Рис, сахарный тростник, бананы, огурец |
|
Анды от Колумбии до севера |
Фасоль, тыква, кукуруза, хлопчатник |
|
Чили |
Картофель, томаты |
Кукуруза, табак, хлопчатник |
В центрах происхождения культурных растений выявлена сопряженная эволюция паразитов и их хозяев. По сообщению Н.И. Вавилова, наибольшее разнообразие видов головни на ржи, например, найдено в юго-западной Азии – центре видообразования этой культуры. Из 10 видов головни, паразитирующих на сорго, большинство найдено только в Африке – родине видов сорго. По мере расселения культурных растений с ними мигрировали и вредные организмы, преимущественно с посевным и посадочным материалом [7]. Выявлена межконтинентальная, внутриконтинентальная миграция вредных организмов, миграция их внутри страны, внутри и между хозяйствами, а также из их природных очагов.
Так, из Америки в Европу мигрировали возбудители фитофтороза картофеля, милдью и оидиума винограда, бактериального ожога плодовых, сорняки – амброзия, горчак розовый, из Европы в Америку – возбудители рака картофеля, кольцевой гнили картофеля, ржавчины смородины и веймутовой сосны, голландской болезни вязов; сорняки – пырей, крапива, бодяк, подорожник.
В пределах континентов широко распространились возбудители рака плодовых (из Англии в Голландию, Польшу, Данию и др. страны), стеблевой и бурой ржавчины злаков (из Северо-восточного Китая в Амурскую область).
Внутри страны произошло распространение пузырчатой головни и фузариоза кукурузы по мере продвижения культуры на север. С Украины в Алтайский и Красноярский края в 1967, 1968 гг. завезена мозаика лука. Широко мигрируют с семенными и посадочными материалами внутри страны возбудители пыльной головни, септориозов, аскохитозов сельскохозяйственных культур, фитофтороза картофеля, которые при благоприятных гидротермических условиях вызывают эпифитотии, обуславливая широкомасштабные применения фунгицидов в течение вегетации.
Кроме миграции вредных организмов из первичных, вторичных и последующих центров возделывания сельскохозяйственных культур, они заселили и заселяют агроэкосистемы из местных целинных и залежных участков, которые служат их резерватами и стартовыми площадками для миграций.
Различают три типа передвижения (расселения) вредных организмов: эмиграцию – выселение с занимаемой территории, иммиграцию – вселение на уже занятую территорию новых особей и миграцию – периодический уход и возвращение вредных организмов на данную территорию.
Выделяют группы вредных организмов по связи их жизненного цикла с разными типами экосистем.
|
Первую группу составляют природно-очаговые вредные организмы. К ним относятся, например, возбудители вирусных болезней костреца безостого, скручивания листьев хлопчатника, желтухи бобов и гороха. Возбудители этих болезней по Е.Н. Павловскому циркулируют в природных очагах по схеме: источник возбудителя инфекции – факторы передачи возбудителя – восприимчивые растения. |
|
Вторую группу составляют вредные организмы, жизненный цикл которых проходит только в агроэкосистемах. Связь их жизненного цикла с естественными экосистемами прервана по мере адаптации к агроэкосистемам. Примерами таких вредных организмов могут служить возбудители твердой головни пшеницы и ячменя, штриховатой мозаики ячменя, пшеничный трипс, просяной комарик. |
|
Третью группу составляют вредные организмы, жизненный цикл которых проходит в естественных и агроэкосистемах. К ним относятся, например, возбудители стеблевой и бурой ржавчины злаков, бронзовости томатов, русской мозаики озимой пшеницы, рапсовый цветоед, саранчовые, хлебные клопы, свекловичная щитоноска, южная свекловичная блошка и др. |



Из 59 проанализированных нами широко распространенных и вредоносных возбудителей болезней сельскохозяйственных культур в Сибири, у 61 % жизненный цикл связан с агроэкосистемами, а у 39 % – с агро- и естественными. Это свидетельствует о том, что для 39 % возбудителей болезней недостаточно проведения мер борьбы только в агроэкосистемах, а следует предусматривать меры по предупреждению их массовой миграции из природных очагов (естественных экосистем).
Особенно это важно в отношении воздушно-капельных и трансмиссивных возбудителей инфекций, против которых применяют пестициды и другие дорогостоящие и экологически небезопасные средства.
Многие фитофаги, переселившиеся на посевы сельскохозяйственных культур в агроэкосистемы, также сохранили связь с естественными экосистемами, проходя на многолетних дикорастущих видах дополнительное питание или зимуя в естественных экосистемах. Из 98 широко распространенных и вредоносных в России фитофагов полевых культур 31 % сохранили устойчивую связь с естественными экосистемами, а остальные 69 % адаптировались к агроэкосистемам, проходя в них все фазы жизненного цикла. Связь с естественными экосистемами может носить у 69 % видов случайный характер, который трудно предусмотреть. В то же время в 31 % случаев меры борьбы с фитофагами в агроэкосистемах следует разрабатывать с учетом их жизненного цикла в естественных экосистемах. Например, исконными местами резервации и размножения азиатской саранчи были плавни рек Кубани, Терека, Маныча, Оки, Десны, откуда она перелетала на посевы сельскохозяйственных культур. Своевременное обнаружение первичных очагов азиатской саранчи и их изоляция имеют первостепенное значение в борьбе с данным фитофагом.
Лес служил в качестве первичной стации для малинного жука, яблонного цветоеда, непарного шелкопряда. Лесные обитатели нематоды адаптировались к почвам пашни, сенокосов и пастбищ. С целинных участков на пашню переселились многие широко распространенные в агроэкосистемах виды щелкунов: посевной, полосатый, темный.
Клопы, вредящие зерновым культурам, часто ограничиваются естественными экосистемами и лишь в благоприятные годы переселяются на посевы сельскохозяйственных культур, где активно размножаются. Однако, энтомофаги клопов не всегда следуют за ними из естественных экосистем в агроэкосистемы, в результате чего связи в системе «клопы – энтомофаги» сдвигаются в пользу клопов – вредителей пшеницы. Та же ситуация отмечена в отношении пшеничного трипса, который переселился с многолетних злаковых трав естественных экосистем в агроэкосистемы пшеничных полей и стал опасным вредителем, а его энтомофаги не последовали за ним на посевы пшеницы. Приведенные примеры свидетельствуют о том, что в агроэкосистемах нередко смещаются трофические связи в системах «фитофаги – энтомофаги» в пользу фитофагов, снижается численность и активность многих энтомофагов, так как они обычно существуют за счет нескольких близких видов насекомых в более равновесных естественных экосистемах.
В целом комплексы вредных организмов в агроэкосистемах представляют собой нередко аналоги естественных экосистем, особенно в отношении наземных видов. Наиболее устойчивыми компонентами, в меньшей степени зависящими от смены сельскохозяйственных культур, являются почвенные вредные организмы. Пастбище – сенокос – сеяные травы – однолетние травы – зерновые – пропашные – таков примерно порядок усиления антропогенного воздействия и постепенной адаптации вредной почвенной фауны естественных экосистем к агроэкосистемам.
Переход из естественных экосистем в агроэкосистемы сопровождается сменой для фитофагов пищи сложного растительного покрова на сравнительно более однообразную. Вследствие этого происходит изменение состава вредных организмов на более специализированные виды.
При освоении целины на культурные злаки перешли с дикорастущих хлебные жуки, хлебные жужелицы, стеблевые блошки, злаковые мухи, вредная черепашка и др. На культурные капустные с дикорастущих крестоцветных растений перешли крестоцветные блошки, капустная белянка, капустная моль, капустная муха и др. На сахарную свеклу с дикорастущих маревых мигрировали свекловичный долгоносик, свекловичная блошка, щитоноска, клопы. Щелкуны, озимая совка, саранчовые стали вредителями многих сельскохозяйственных культур. Формирование вредной фауны плодовых культур происходило путем перехода на них насекомых с боярышника, рябины, черемухи, диких плодовых деревьев и кустарников.
В естественных экосистемах насекомые-фитофаги не наносят растениям невосполнимого урона. В агроэкосистемах численность вредных организмов резко возросла вследствие массового размножения при обилии пищи, уменьшении численности энтомофагов, и они причиняют значительный вред. Однако число вредных видов среди фитофагов не превышает 1–2 %. Детальное изучение фауны озимой пшеницы в лесостепной зоне показало, что только 1,5 % видов питаются пшеницей. Остальные виды составляют зоофаги, некрофаги, капрофаги, сапрофаги. Зоофаги, составляющие 70 % от общей фауны, активизируются только к уборке урожая пшеницы, а поэтому не оказывают влияние на процессы саморегуляции видов в агроэкосистемах.
|
В целом сукцессионные процессы в агроэкосистемах протекают быстрее, чем процессы адаптации к ним популяций вредных организмов. В этой связи менее пластичные виды элиминируются (вытесняются), в том числе расселяются на другие стации. Замечено, что чем уже экологический диапазон адаптации популяций вида, тем важнее для него функция расселения. |

Из форм с многолетним жизненным циклом в агроэкосистемах выживают только многоядные виды (полифаги) в личиночной стадии (проволочники, ложнопроволочники, личинки пластинчатоусых) зачастую обладая способностью к сапрофагии. Из олигофагов хорошо адаптировались к агроэкосистемам виды с коротким (не более одного года) жизненным циклом и высокой способностью к расселению. Примером изменения сообщества фитофагов в агроэкосистемах по сравнению с естественными (целинные участки) могут служить данные табл. 19.
Таблица 19
Изменения сообщества фитофагов при распашке целины в Оренбургских степях [2]
Показатели |
Целинная степь |
Пшеничное поле |
Число видов насекомых, в т.ч.: |
312 |
135 |
цикадовые |
35 |
12 |
клопы |
38 |
19 |
жуки |
93 |
39 |
перепончатокрылые |
37 |
18 |
Обилие (число особей) на 1 кв.м. |
159 |
341 |
Число доминантных видов |
38 |
19 |
Их общее обилие на 1 кв.м. |
106 |
332 |
Фауна насекомых в естественных экосистемах (целина) примерно в два раза богаче, чем в агроэкосистемах (посев яровой пшеницы). Однако численность фитофагов на пшеничном поле в два раза выше, чем на целинном участке. Следовательно, освоение целины в Оренбургской области привело к элиминации (гибели) 37 % видов, но выжившие виды – хлебная полосатая блошка, серая зерновая совка, пшеничный трипс и др. – стали более многочисленны (их численность возросла в 20–280 раз).
Вредителями полевых культур стали олигофаги, которые в личиночной стадии малоподвижны и обладают ограниченной способностью к активному расселению в стадии имаго. Эти особенности компенсировались их пластичностью и специализацией к растениям в агроэкосистемах.
В целом агроэкосистемы имеют меньшее видовое разнообразие организмов, чем естественные. Огород, поле, сад беднее видами, чем лес, луг, степная ассоциация. При этом в любой экосистеме можно выделить один или несколько доминирующих видов. Для оценки количественного представления видов в популяциях используют индекс разнообразия, который вычисляется по формуле Шеннона:

где: 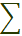 – знак суммы
– знак суммы
Pi – доля каждого вида в сообществе (по численности или массе)
log2Рi – двоичный логарифм Pi.
Для оценки роли отдельных видов в структуре сообщества определяют следующие показатели:
 обилие вида – число особей вида на единице площади или объеме занимаемого пространства. Для оценки используют 5-бальную шкалу в баллах:
обилие вида – число особей вида на единице площади или объеме занимаемого пространства. Для оценки используют 5-бальную шкалу в баллах:
0 – отсутствует;
1 – редко и рассеянно встречается;
2 – нередко встречается;
3 – обильно встречается;
4 – очень обильно встречается (доминирует);
 частоту встречаемости – характеризует равномерность или неравномерность распределения популяции вида в сообществе. Ее рассчитывают как процентное отношение числа проб, где встречается вид;
частоту встречаемости – характеризует равномерность или неравномерность распределения популяции вида в сообществе. Ее рассчитывают как процентное отношение числа проб, где встречается вид;
 постоянство вида выражают по формуле:
постоянство вида выражают по формуле:
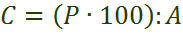
где: Р – число выборок, содержащих вид
А – общее число выборок.
Для оценки постоянства видов применяется шкала:
постоянные виды |
– |
более 50 % выборки |
добавочные |
– |
25–50 % |
случайные |
– |
менее 25 % |
 доминирующие виды, занимающие господствующее положение, определяют по степени доминирования – показателю числа особей данного вида к числу всех особей в %.
доминирующие виды, занимающие господствующее положение, определяют по степени доминирования – показателю числа особей данного вида к числу всех особей в %.
Сообщество вредных организмов в агроэкосистемах формируется вокруг растений-хозяев, которые выступают в качестве эдификаторов-доминантов, или центральных видов в сообществах, определяющих их особенность. Например, сообщество вредных организмов яровой пшеницы в северной лесостепи Приобья Западной Сибири насчитывает более 40 распространенных и вредоносных видовых популяций. Вместе с энтомофагами, антагонистами, микроорганизмами ризосферы и филлопланы, эпифитами и другими видами они составляют своеобразную структуру – консорцию, присущую пшеничному полю.
Под консорцией понимают сообщество организмов, формирующееся вокруг эдификаторов, жизнедеятельность которых трофически или топически связана с эдификатором – автотрофным растением. |
Среди консортов есть виды, входящие в состав только одной консорции, являющиеся членами двух и более консорций, образуя взаимосвязанные комплексы. Связи между видовыми популяциями в консорции могут быть длительные (на протяжении всей жизни) или носят сезонный характер, обусловленный адаптацией их к определенным экологическим условиям. Виды в консорциях выполняют различные функции в экосистемах и связаны между собой различными взаимоотношениями (нейтрализм, конкуренция, паразитизм, хищничество и др.).
Сорные растения являются в основном свободноживущими. Казанские фитоценологи выяснили, что в агроэкосистемах озимой ржи сорняки были представлены видами, из которых 83,3 % произрастали и в других экосистемах сельскохозяйственных культур. В агроэкосистемах яровых зерновых культур такие виды сорняков составляли 93,0 %. По данным крымских фитоценологов, независимые виды сорняков (свободноживущие) преобладали в 52,0–79,0 % случаев. Тем не менее, выявлены отдельные виды сорняков, тяготеющие к определенным сельскохозяйственным культурам: к яровой пшенице – сурепка, к кукурузе – горец шершавый, к овсу – овсюг.
|
В зависимости от географического положения агроэкосистем, сообществам вреднях организмов, как и других биологических видов, присущи характерные особенности. Мягкому климату соответствует разнообразие и обилие видовых форм жизни, усложнение сообществ, специализация видов и сужение их экологических ниш. Этому сопутствует устойчивая численность и высокая степень оседлости видов. Естественный отбор в таких условиях способствует формированию видов с признаками К-стратегов, то есть носит стабилизирующий характер. |

Возрастание суровости и изменчивости климата в умеренных и высоких широтах сопровождается уменьшением видового разнообразия, биомассы и упрощением строения сообществ. Это совпадает с приобретением в процессе эволюции под давлением направленного (движущего) отбора признаков r-стратегов, появлению и усилению роли сезонного изменения сообществ одновременно с уменьшением суточного изменения, полностью исчезающего в высоких широтах. Нарастает подвижность животных, переходящих ко все более дальним сезонным миграциям. Одновременно увеличивается неустойчивость численности отдельных видов и изменчивость облика биоценозов в целом, в связи с чем, меняется сила и характер связей между видами. Устойчивые межвидовые связи тропических биоценозов превращаются в умеренных и высоких широтах в подвижные и непостоянные. Цепи питания то возникают, то распадаются, и все сообщество становится изменчивым по составу и структуре. Крайней степени эти особенности достигают в открытых ландшафтах с несложными, часто одноярусными сообществами (тундра, степь). В связи с этим относительная роль биотических условий (враги, паразиты, конкуренты) падает, и на первый план выступают абиотические условия, прямо или косвенно определяющие исход взаимоотношений в сообществах (рис. 6).
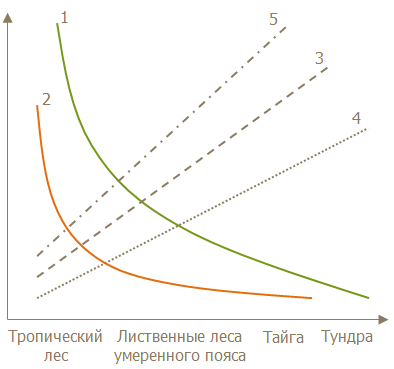
Рис. 6. Схема изменения структуры сообществ в различных зонах [23]
1 – общее число видов; 2 – доля видов, ведущих ночной образ жизни; 3 – доля видов, запасающих корм; 4 – доля видов-мигрантов; 5 – степень изменчивости численности
В процессе переселения вредные организмы, особенно фитофаги, проникают в новые для них географические ареалы. Возбудители болезней в большей мере космополиты, в то время как фитофаги приурочены к определенным географическим зонам, характеризующимся определенными гидротермическими условиями. Поэтому знание зональности видов в защите растений очень важно.
Территория, на (в) которой распространен вид, независимо от его численности называется ареалом вида. |
Часть ареала, на которой фитофаг часто встречается в значительном количестве и может причинять большие повреждения сельскохозяйственным культурам, называется зоной вредности. |
В зоне вредности фитофаг также распространен очень неравномерно, а приурочен к определенным стациям, участкам с определенными экологическими и хозяйственными условиями. Например, сибирская кобылка развивается на залежных и пастбищных участках с плотной почвой, благоприятной для откладки яиц в кубышки по 7–10 штук. Самка сибирской кобылки откладывает в этих стациях яйца в 9–18 кубышек. В то же время личинки и имаго предпочитают другие стации – посевы пшеницы, ячменя, а из многолетних злаков – пырея, костреца.
Стации одного и того же вида изменяются в различных географических зонах. Так, азиатская саранча на юге заселяет плавни рек, а в средней полосе – открытые, хорошо прогреваемые песчаные участки, которые по температурным условиям приближаются к плавням южных рек.
|
В общей сложности выделяют два типа географических ареалов вредных организмов: глобальные и региональные. При глобальных ареалах вредные организмы распространены повсеместно (ржавчинные заболевания, обыкновенная корневая гниль хлебных злаков), а при региональных – ограничены определенными областями (гречиха татарская, жук-кузька). |

Суточная периодичность, свойственная насекомым и возбудителям болезней, приводит к формированию «дневных» и «ночных» видов, которые являются составной частью двух, до некоторой степени самостоятельных, сменяющих друг друга, суточных сообществ. Этот путь вел к усложнению сообществ и более полному использованию ресурсов среды.
Сезонные изменения сообществ зависят от впадения отдельных видов в летнюю или зимнюю спячку, миграций при наступлении засухи или зимы. Адаптированные к агроэкосистемам оседлые виды (их доля превышает 60 % среди широко распространенных и вредоносных видов) составляют ядро сообществ в агроэкосистемах.
Эволюционные изменения популяций отдельных видов и сообществ находятся в тесном взаимодействии. Изменение среды меняет направление естественного отбора, тактик жизнедеятельности и стратегию жизненных циклов видов, а это в свою очередь изменяет взаимоотношения видов в сообществах, их экологические ниши. Эти процессы хотя и сопряжены, но все же независимы, так как изменение сообществ может происходить и без эволюции входящих в них видов, а появление нового вида не обязательно изменяет тип сообщества. Это положение важно для понимания структуры сообществ в агроэкосистемах, которые изменяются в результате миграций более 30 % особо вредоносных организмов, а также изменением технологий возделывания сельскохозяйственных культур с целью более полной их адаптации к почвенно-климатическим и экономическим условиям.
Таким образом, эволюционно-экологическое рассмотрение закономерностей формирования сообществ вредных организмов в агроэкосистемах свидетельствует о различной степени адаптации отдельных видов. Часть (более 30 %) видов сохранили генетическую память своей связи с естественными экосистемами, устойчиво мигрируя туда в процессе жизненного цикла, особенно для перезимовки и дополнительного питания. Некоторые виды формируют в естественных экосистемах природные очаги, которые представляют собой стартовые площадки для переселения в агроэкосистемы. Однако, большинство вредных организмов (более 60 %) адаптировано к прохождению всех фаз жизненного цикла в агроэкосистемах, формируя ядро сообществ вредных организмов возделываемых культур. Все виды и сообщества вредных организмов в целом подвержены естественному отбору, активаторами или депрессантами которого служат природные (климатические) и антропогенные (технологии возделывания) факторы. Это приводит к формированию сообществ, в которых преобладают виды с признаками r- или К-стратегов, а также виды со смешанными признаками. В агроэкосистемах естественный отбор в большей степени благоприятствует формированию признаков r-стратегов, то есть носит направленный (движущий) характер. Особенно это характерно для наземно-воздушных вредных организмов в зонах с неустойчивым континентальным климатом.
Сообщества приобретают изменчивый характер, обусловленный входящими в его состав доминирующими видами с признаками r-стратегов со значительной зависимостью их численности от абиотических и антропогенных факторов.


