 Экологические основы интегрированной защиты растений: учебник
Экологические основы интегрированной защиты растений: учебник

 ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ

 ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ


6.2. Эпифитотиологические группы вредных организмов в агроэкосистемах
Закономерности формирования сообществ вредных организмов в агроэкосистемах изложены в ГЛАВЕ 3.
Сообщества вредных организмов в агроэкосистемах формируются вокруг растений сельскохозяйственной культуры, которая является эдификатором-доминантом, или средообразующим видом в агроценозе. Исследования показали, что сообщества опасных вредных организмов на одной культуре состоят в среднем из 44 видов (варьирование от 29 до 60), которые принадлежат к трем группам экологических эквивалентов: почвенным, или корне-клубневым, наземно-воздушным, или листо-стеблевым и трансмиссивным. Семенные вредные организмы входят в состав сообществ только на 58,8 % культур. На остальных 41,2 % культур они отсутствуют, хотя семена могут служить дополнительным фактором передачи вредных организмов. Например, гречиха, рапс, лен, конопля не имеют специализированных семенных инфекций, у которых возбудители передавались бы во времени от маточных растений к дочерним только через семена. В то же время семена всех этих культур могут поражаться возбудителями из других эпифитотиологических групп, например, видами рода Fusarium, которые принадлежат к почвенным, или корне-клубневым, вредным организмам.
|
Для разработки систем ИЗР на уровне севооборотов и агроландшафтов чрезвычайно важны знания связи жизненного цикла вредных организмов с разными типами экосистем. По данному критерию все вредные организмы разделены на два класса (группы). |


К первой группе относятся вредные организмы, жизненный цикл которых проходит в естественных и агроэкосистемах. Эволюционно-экологические тактики Р, В, Т таких вредных организмов сохранили внутреннюю эволюционную стабильность, или (в терминах эволюционной экологии) – эволюционно стабильную стратегию (ЭСС) поведения (реакции) и саморегуляции хотя степень стабильности может существенно различаться в природных и агроэкосистемах. Свойства жизненного цикла всех вредных организмов указанной группы могут служить эталоном для оценки изменения тактик Р, В, Т и стратегии жизненного цикла вредных организмов в агроэкосистемах, а следовательно, и для разработки систем ИЗР с ними.
Эта группа вредных организмов недостаточно изучена. Первые исследования свидетельствуют о том, что видовые популяции естественных (природных) экосистем отличаются значительно меньшей интенсивностью размножения по сравнению с агроэкосистемами. Кроме того, их тактика выживания лучше адаптирована к условиям окружающей среды. У фитопатогенов, например у Botrytis cinerea, в естественных экосистемах формируются более крупные склероции, а у возбудителей твердой головни телиоспоры сохраняются сорусами, а не в распыленном состоянии, как в агроэкосистемах.
Из естественных экосистем вредные организмы переселяются в агроэкосистемы. По этой причине состав вредных организмов агроэкосистем по видовому составу нередко близок к естественным экосистемам.
В естественных экосистемах на многолетних злаках (тимофеевка луговая, пырей ползучий, овсяница луговая, регнерия волокнистая и др.) постоянно паразитирует возбудители корневых гнилей яровой пшеницы и ячменя Bipolaris sorokiniana, а также виды рода Fusarium (F. sporotrichioides, F. poae, F. culmorum, F. equiseti, F.oxysporum var. orthoceras и др.) – представители почвенных или корне-клубневых, вредных организмов. Из трансмиссивных инфекций к числу природно-очаговых относятся возбудители вирусных болезней костреца безостого, скручивания листьев хлопчатника, желтухи бобов и гороха и др.
Фитофаги, переселившиеся на посевы сельскохозяйственных культур в агроэкосистемы, сохранили генетическую память связи жизненного цикла с естественными экосистемами, проходя на многолетних дикорастущих видах дополнительное питание или зимуя в естественных экосистемах. Из 98 широко распространенных и вредоносных в России фитофагов полевых культур 30 % сохранили устойчивую связь с естественными экосистемами. Например, исконными местами резервации и размножения азиатской саранчи на Юге России являются плавни рек Кубани, Терека, Манычи и других рек, откуда они перелетают на посевы сельскохозяйственных культур. Своевременное обнаружение первичных очагов азиатской саранчи и их локализация имеют первостепенное значение в борьбе с данным фитофагом.
Сорные растения, относящиеся к свободноживущим видам, приурочены как к естественным, так и агроэкосистемам. Только у некоторых видов сорняков жизненный цикл тяготеет к определенным сельскохозяйственным культурам в агроэкосистемах: ко льну – плевел льняной, к кукурузе – горец шершавый, к овсу – овсюг.
Частота переходов, и соответственно генетических обменов, популяций вредных организмов из природных в агроэкосистемы и обратно изучена недостаточно, хотя имеет несомненное практическое значение в плане разработки систем мониторинга, прогноза и защитных мероприятий. По-видимому, можно полагать, что частота переходов вредных организмов между различными типами экосистем определяется их миграционными способностями и выше у групп листо-стеблевых и трансмиссивных (r- и rK-стратегов), чем у семенных, а особенно почвенных видов (Кr- и К-стратегов).

Ко второй группе относятся вредные организмы, жизненный цикл которых проходит преимущественно в агроэкосистемах. Связь их жизненного цикла с естественными экосистемами практически прервана. Тактики Р, В, Т таких вредных организмов нарушены в процессе хозяйственной деятельности человека. В их жизненном цикле нередко «выпадение» некоторых фаз (стадий развития).
Анализ жизненного цикла 157 фитопатогенов и фитофагов позволил выяснить, что примерно у 60 % фитопатогенов и 70 % фитофагов жизненный цикл адаптирован преимущественно к агроэкосистемам, а связь с естественными экосистемами носит случайный характер. Например, лес служил в качестве первичной стации для малинного жука, яблонного цветоеда. В настоящее время эти виды обитают преимущественно в агроэкосистемах. Лесные нематоды адаптировались к почвам пашни, сенокосов и пастбищ. С целинных участков на пашню переселились многие широко распространенные в агроэкосистемах виды щелкунов: посевной, полосатый, темный.
Примером фитопатогенов, жизненный цикл которых адаптирован преимущественно к агроэкосистемам, могут служить бугорчатая парша и фитофтороз картофеля, твердая головня пшеницы.
Системы ИЗР против вредных организмов рассматриваемой группы разрабатываются преимущественно первых четырех уровней сложности – против популяций, групп экологических эквивалентов, сообществ культур и севооборотов.
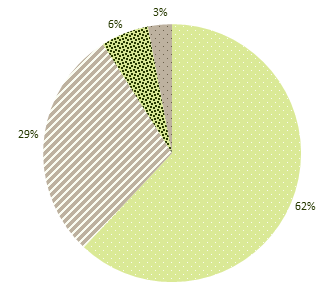
Усредненная структура сообществ наиболее распространенных вредных организмов сельскохозяйственных культур в полевых севооборотах представлена на рис. 34.
|
|
|
|
Рис. 34. Структура сообщества вредных организмов (среднее по 17 сельскохозяйственным культурам)
Доминирующая роль в структуре сообществ вредных организмов по культурам в севооборотах принадлежит наземным, или листо-стеблевым видам. Их доля изменяется по разным культурам от 51,4 % (зернобобовые культуры) до 72,7 %. Исключение составляет лишь картофель, по которому доля наземно-воздушных, или листо-стеблевых, вредных организмов составляет 34,6 %.
Второе место в структуре сообществ вредных организмов принадлежит почвенным, или корне-клубневым видам. Их доля изменяется от 11,4 % (рис) до 50,0 % (картофель), а в среднем составляет 28,9 %, или в 2,1 раза меньше, чем наземно-воздушных, или листо-стеблевых вредных организмов.
Во всех зонах численность вредных организмов в различных эпифитотиологических группах в агроэкосистемах намного выше, чем в естественных. Покажем это на примере широко распространенного основного возбудителя корневых гнилей хлебных злаков (табл. 53).
Таблица 53
Фитосанитарное состояние почв разного типа в агро- и естественных экосистемах Западной Сибири
Зона, экосистема |
Тип почвы |
Численность конидий B.sorokiniana в 1г почвы |
Рост по сравнению с естественной экосистемой, число раз |
Подтайга |
|||
Агроэкосистема |
Темно-серая |
30±3,0 |
6,0 |
Естественная экосистема |
Темно-серая |
5,0±1,7 |
|
Естественная экосистема |
Луговая северная |
1,7±0,2 |
|
Предгорья |
|||
Агроэкосистема |
Чернозем выщелоченный |
141±0,2 |
|
Северная лесостепь |
|||
Агроэкосистема |
Чернозем выщелоченный |
89,0±2,0 |
8,1 |
Естественная экосистема |
Чернозем выщелоченный |
11,0±1,7 |
|
Южная лесостепь |
|||
Агроэкосистема |
Чернозем выщелоченный |
84,0±5,0 |
16,8 |
Естественная экосистема |
Чернозем выщелоченный |
5,0±1,7 |
|
Естественная экосистема |
Чернозем обыкновенный |
13,3±0,1 |
|
Агроэкосистема |
Темно-каштановая |
5,8,0±4,0 |
5,8 |
Естественная экосистема |
Темно-каштановая |
10,0±2,0 |
|
Степь |
|||
Агроэкосистема |
Южный чернозем |
73,0±3,0 |
6,2 |
Естественная экосистема |
Южный чернозем |
11,7±1,2 |
|
Среднее |
|||
Агроэкосистема |
Паровое поле |
79,2±1,7 |
9,7 |
Естественная экосистема |
Фитоценозы со злаками |
8,2±0,9 |
|
Самая высокая заселенность почвы возбудителем корневых гнилей отмечена в предгорьях южной и северной лесостепи, где складываются благоприятные условия не только для возделывания ячменя и яровой пшеницы, но и для интенсивной споруляции B.sorokiniana на прикорневых листьях растений.
На примере болезней картофеля различной эпифитотиологии было выяснено, что почвенные, или корне-клубневые, вредные организмы приурочены к северным агроландшафтам, а надземные, или листо-стеблевые, к увлажненным южным агроландшафтам лесостепи и предгорий.
Подобные закономерности И.Ф. Павлов отмечает в распределении фитофагов. В северных широтах поливольтинные виды фитофагов дают 1–2 поколения, в то время как в южных – 3–4 и более. В тундре, где крайне малочисленный видовой состав вредных организмов, отмечена высокая вредоносность личинок (проволочников) темного щелкуна.
Во всех зонах популяции вредных организмов в большей степени подвержены антропогенному воздействию в пределах агроценозов (севооборотов), чем за их пределами в агроландшафтах, особенно на нераспаханных землях. При этом естественный отбор в агроценозах благоприятствует выработке у вредных организмов признаков r-стратегов.


