| Начало темы |
21.1.3. Селезёнка
21.1.3.1. Основные компоненты селезёнки
В селезёнке можно
выделить 4 основные компонента:
|
I. Капсула и трабекулы
| 1. Трабекулы
(2)
|
Схема -
строение селезёнки.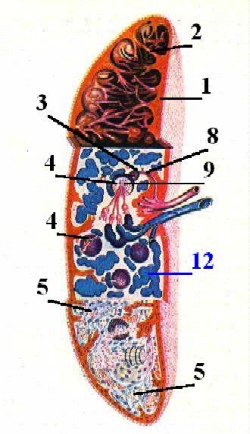 |
II. Белая пульпа
1. Белая пульпа - лимфоидная ткань, включающая
проникших в кровь через барьер лимфоузлов. |
III. Красная пульпа
| 1. Красная пульпа - содержимое пространства между лимфоидной тканью и трабекулами. 2. Её компоненты таковы. -
3. а) Макрофаги в селезёночных тяжах захватывают и разрушают старые эритроциты и тромбоциты. б) А в синусах может депонироваться определённое количество крови. |
IV. Специфическая сосудистая система селезёнки
Перечень сосудов
| Сосуды данной
системы
|
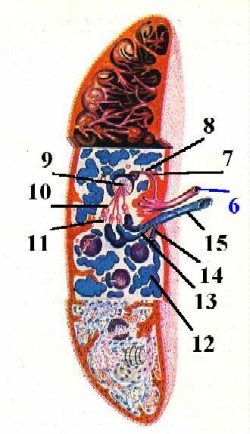 |
Особенности сосудистой системы селезёнки
Укажем две главные особенности этой системы. -
| Специфи- ческие образования в стенке сосудов |
В стенке ряда сосудов
имеются специфические образования -
|
| Наличие
открытого кровообра- щения |
1. А среди капилляров
2. Во втором случае
3. При переполнении синусов происходит обратное:
|
21.1.3.2. Функции и развитие селезёнки
I. Функции
| Из вышеизложенного
следуют функции селезёнки. – 1. Депонирование крови (в красной пульпе) и отдельных её элементов (тромбоцитов, эритроцитов), а при необходимости (напр., после кровопотери) -
2. Элиминация (удаление из кровотока и разрушение) старых и повреждённых эритроцитов и тромбоцитов. (Это опять
связано с красной пульпой.) 3. Участие в иммунной защите от чужеродных антигенов. - Обеспечивается
|
II. Развитие
| Развитие селезёнки
отражается схемой. –
|
Теперь подробнее рассмотрим строение селезёнки взрослого.
21.1.3.3. Опорно-двигательный аппарат селезёнки
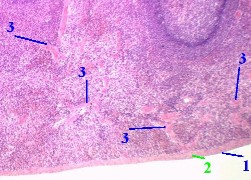
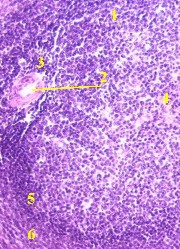
| 7,а-б. Препарат - селезёнка; окраска гематоксилин-эозином. | |
1. С
поверхности селезёнка покрыта серозной оболочкой, включающей
2. Глубже располагается капсула (2), образованная, как отмечалось, плотной волокнистой соединительной тканью с гладкими миоцитами. 3. а) На снимках мы видим также фрагменты многочисленных трабекул (3), образованных той же тканью и гладкими миоцитами. б) Их можно узнать по
|
|
21.1.3.4. Белая пульпа
I. Четыре зоны лимфатического узелка
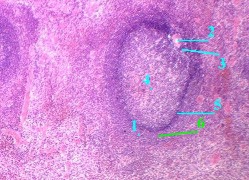
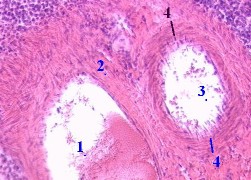
| 7,в-г. Препарат - селезёнка; окраска гематоксилин-эозином. | |||||||||
| 1. Здесь в поле зрения - один из
лимфоидных узелков, или фолликулов (1) селезёнки. 2. На периферии его мы видим артерию узелка, или т.н.
центральную артерию (2), 3. На срезе узелка выделяют несколько зон. Это:
|
|||||||||
II. Функция и клеточный состав
а) По своей функции и клеточному составу зоны фолликулов селезёнки аналогичны соответствующим образованиям лимфоузлов.
б) Учитывая ещё селезёночные тяжи красной пульпы (п. 21.1.3.1.II), получаем следующую таблицу (ср. с таблицей в п. 21.1.1.5).
| СТРУКТУРА СЕЛЕЗЁНКИ |
АНАЛОГИЧНАЯ СТРУКТУРА ЛИМФОУЗЛА |
К Л Е Т К И (ПОМИМО РЕТИКУЛЯРНЫХ) |
| ПЕРИ- АРТЕРИАЛЬНАЯ ЗОНА |
ПАРА- КОРТИКАЛЬНАЯ ЗОНА |
1. Т-лимфоциты
на разной стадии антигензависимой
дифференцировки. 2. Интердигитирующие клетки: |
| ГЕРМИНАТИВНЫЙ
ЦЕНТР (реактивная зона) |
ГЕРМИНАТИВНЫЙ
ЦЕНТР (реактивная зона) |
1. В-иммунобласты. 2. Дендритные клетки: длительно сохраняют на поверхности антигены. 3. Крупные макрофаги. |
| МАНТИЙНАЯ
ЗОНА |
"КОРОНА" ФОЛЛИКУЛА | 1. Малые В-лимфоциты -
2. Макрофаги. |
| КРАЕВАЯ,
ИЛИ МАРГИНАЛЬНАЯ ЗОНА |
------------- | В- и
Т-лимфоциты.
|
| СЕЛЕЗЁНОЧНЫЕ ТЯЖИ КРАСНОЙ ПУЛЬПЫ | МОЗГОВЫЕ
ТЯЖИ ЛИМФОУЗЛОВ |
1.
Проплазмоциты и плазматические клетки. 2. Макрофаги. 3. В селезёночных тяжах - ещё элементы крови (поступающие из капилляров с открытыми концами). |
21.1.3.5. Красная пульпа
I. Просмотр препарата
II. Селезёночные тяжи:
плазматические клетки
| Плазмоцит
из селезёнки. Электронная микрофотография. 1. а) Согласно п. 21.1.3.4.II, важным компонентом селезёночных тяжей являются плазматические клетки. б) Здесь приведена микрофотография одной из них. 2. а) В связи с интенсивным синтезом экспортных белков (иммуноглобулинов), в этих клетках хорошо развиты |
|
| б) Причём, последний |
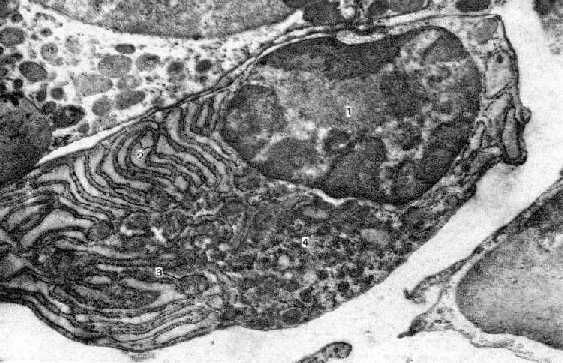 |
3. В клетке также можно видеть митохондрии (3). |
|
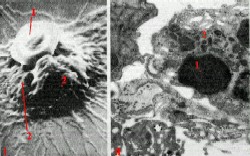
III. Селезёночные тяжи: разрушение эритроцитов
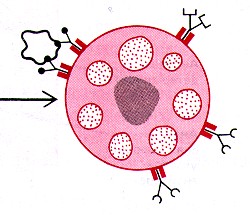
Узнавание старых эритроцитов макрофагами
| Электронные
микрофотографии - фагоцитоз эритроцитов в селезёнке : I - сканирующая и |
|
1. Как отмечалось, в селезёночных тяжах старые и повреждённые эритроциты (1) захватываются и разрушаются макрофагами (2). 2. а) По мере старения эритроциты постепенно теряют отрицательно заряженные остатки сиаловой кислоты на своей поверхности. б) Видимо, снижение заряда до критического значения и является тем признаком, по которому узнаются "старые" эритроциты. 3. Кроме того, может иметь значение и то, что
|
|
Продукты распада гемоглобина
Судьба гемоглобина фагоцитированных эритроцитов отражается схемой. –
|
| Образова- ние желчных пигментов |
1. Желчные пигменты (билирубин), согласно схеме - продукты распада небелковой части гемоглобина.
2. Они
3. После этого
4. В результате, именно с этими пигментами связан жёлтый цвет кала и мочи. |
| Реутилиза- ция железа |
А ионы железа, высвобождающиеся в селезёнке при
распаде гемоглобина, большей частью реутилизируются:
|
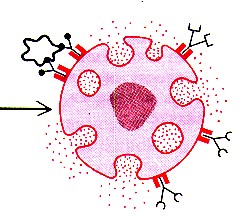
IV. Венозные синусы
| Общие особенности | Венозные
синусы в селезёнке имеют три особенности,
общие для
микрососудов подобного типа (п. 18.3.1.2):
|
|
| Специфичес- кие особенности строения |
Кроме того,
существуют и специфические
особенности:
|
|
| Функциональ- ные особенности |
Отсюда
вытекают упоминавшиеся выше два свойства
синусов селезёнки: а) они способны депонировать кровь, б) через их стенку могут проходить элементы крови:
|
|
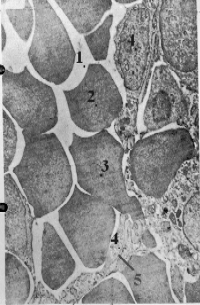
| Иллюстра- ция |
Вторая ситуация
представлена на снимке. -
|
 |
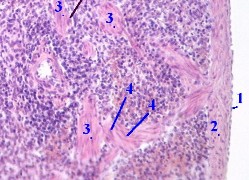
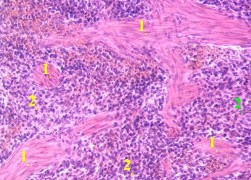
21.1.3.6. Сосуды трабекул
| 1. а) Обратим внимание на
особенность других сосудов селезёнки - трабекулярных вен (1). – б) Они относятся к венам безмышечного типа (п. 19.1.2) 2. Поэтому в этих венах имеются лишь
3. В результате, вены
селезёнки |
7,е. Препарат - селезёнка; окраска гематоксилин-эозином. |
| 4. Трабекулярные артерии (3) отличаются от одноимённых вен наличием циркулярного слоя гладких миоцитов в средней оболочке (4). | |
![]()
Как мы знаем, функция
лимфоидной ткани, содержащейся в рассмотренных
выше органах, - это участие в иммунных процессах.
Остановимся на данной функции немного подробнее.
21.2. Краткие сведения об иммунных процессах
Иммунные процессы - это процессы, защищающие организм от частиц (вирусов, бактерий, различных клеток) и растворимых веществ, несущих на себе признаки генетической чужеродности.
Иными словами, это процессы распознавания "чужого" и защиты от него.
21.2.1. Основные участники иммунных процессов
21.2.1.1. Антигены
| Полные антигены 1. Вещества, которые в отдельном (растворённом) виде или в составе частиц (вирусов, бактерий, клеток)
называются антигенами (или полными антигенами). 2. Антигенностью обладают различные соединения с достаточно высокой молекулярной массой - чужеродные
|
| Гаптены. 1. Некоторые вещества, называемые гаптенами, вызывают специфическую иммунную реакцию
2. Гаптенами являются чужеродные
|
| Антигенные
детерминанты. 1. Те части антигена или гаптена, которые определяют специфичность иммунной реакции, называются антигенными детерминантами. 2. Один антиген (например, чужеродный белок)
может иметь несколько
разных антигенных детерминант - 3. В свою
очередь, частица (вирус, бактерия, клетка) может
иметь много
разных антигенов. 4. Поэтому в ответ на попадание в организм какого-либо микроорганизма может образовываться сразу много разных антител. |
21.2.1.2. Главный комплекс гистосовместимости
I. Определения
1. Очень
важную роль во многих иммунных реакциях
играют т.н. антигены
главного комплекса гистосовместимости (антигены ГКГ).
2. Различают 2 класса данных антигенов. |
II. Характеристика антигенов ГКГ
| Антигены ГКГ класса I. 1. а) Весь набор этих антигенов (по разным оценкам - от нескольких десятков до нескольких сотен) содержится
б) Здесь на него приходится ~ 1% белков плазмолеммы (~ 500.000 молекул на клетку).
б) Поэтому клетки разных людей
отличаются по набору антигенов ГКГ-I, 3. Гены ГКГ-I у человека локализуются в 6-й хромосоме. 4. Многие наследственные болезни иммунного характера связаны с наличием определённой аллели какого-либо из генов ГКГ-I. |
| Антигены ГКГ класса
II. Эти антигены имеются на поверхности лишь клеток, участвующих в представлении антигенов, а именно на поверхности
|
III. Роль антигенов ГКГ в
иммунных процессах
| 1. Данные гликопротеины - это
своего рода маяки, сигнализирующие об отсутствии
или наличии на поверхности клетки "чужого". - а) Если все антигены ГКГ ни с чем (кроме самой клетки) не связаны и не изменены,
б) Если же какой-либо из антигенов ГКГ неспецифически связал чужеродный антиген или как-то изменился,
|
| 2. В частности, а) Т-киллеры атакуют комплексы вида
б) а Т-хелперы - комплексы
|
21.2.1.3. Клоны В- и Т-клеток
I. Иммуноспецифичность клонов
| 1. Со стороны
организма основными участниками иммунных
процессов являются, как мы знаем, В- и Т-лимфоциты. 2. Мы уже также отмечали (п. 20.2.2.4.II), что на начальных этапах созревания лимфоцитов происходит
образующиеся клетки различаются по гену, кодирующему иммунорецепторы и иммуноглобулины. 3. Потомки этих клеток формируют клоны (примерно 107), каждый из которых
4. В то же время, одна антигенная детерминанта иногда способна реагировать (с большим или меньшим сродством) с рецепторами нескольких разных клонов - и, в результате, активировать сразу все эти клоны. |
II. Гипотетический механизм образования генов Ig
| Образование новых генов
при созревании лимфоцитов происходит, видимо,
следующим образом. - 1. В исходных клетках имеется всего несколько сот коротких генов, кодирующих фрагменты пептидных цепей будущих антител: примерно
2. Затем же (на определённой стадии созревания) эти первичные гены случайным образом объединяются друг с другом, формируя полные гены иммуноглобулинов. 3. Из-за случайного характера этого процесса, в разных клетках получаются
4. Расчёт
показывает: |
21.2.1.4. Макрофаги
| Виды макрофагов |
а) Важный участник многих иммунных
процессов -
б) Они часто участвуют
|
| Роль в индуктивной фазе | В начальной фазе имеет значение то, что макрофаги
|
| Роль в
продуктив- ной фазе |
В конечной же фазе иммунной реакции макрофаги интенсивно фагоцитируют продукты взаимодействия антител с антигенами:
|
21.2.1.5. Гранулоцитарные лейкоциты и тучные клетки
| Нейтрофилы | а) А. Многие нейтрофилы тоже
(как и макрофаги) содержат на поверхности рецепторы к FC-области IgG. В связи с этим, они принимают активное участие в
Б. То, какие клетки (макрофаги или нейтрофилы) преимущественно фагоцитируют бактерии в том или ином конкретном случае, зависит от локализации процесса и природы бактерий. б) Однако в отличие от макрофагов, нейтрофилы
|
| Базофилы
и тучные клетки |
а) На поверхности базофилов и тучных
клеток имеются рецепторы к FC-области
иммуноглобулинов класса Е (IgЕ). Соответственно, эти Ig обычно сорбированы на данных клетках. б) При дополнительном же связывании антигена инициируется высвобождение содержимого базофильных гранул - в т.ч. гистамина, который
Т.о., в области нахождения антигена развивается (при наличии Ig E) воспалительная реакция. в) Другие компоненты базофильных гранул, возможно, оказывают
|
| Эозинофилы | а) Повышение в ткани концентрации
гистамина привлекает сюда (по принципу
положительного хемотаксиса) эозинофилы, которые
б) Ключевую роль играют эозинофилы в реакции на паразиты: эта реакция (видимо, тоже включающая образование антител) заканчивается
|
21.2.2. Иммунные реакции
Различают 2 основных типа иммунных реакций:
клеточный и
21.2.2.1. Общая характеристика
| Клеточная иммунная реакция |
Гуморальная иммунная реакция |
|
| Агенты, вызывающие реакцию |
Антигенными агентами служат эукариотические клетки с
"чужеродными" антигенами ГКГ класса I на
поверхности (п. 21.2.1.2):
|
Гуморальную реакцию вызывают:
а также достаточно мелкие корпускулярные антигены:
|
| Суть реакции |
Указанные клетки атакуются и
разрушаются |
Антиген атакуется специфическими
и образующиеся комплексы фагоцитируются макрофагами или нейтрофилами. |
| Клетки, участвующие в реакции | Среди киллеров (п. 20.2.5.4) отметим
|
В гуморальной реакции круг
вовлечённых клеток шире:
|
|
||
21.2.2.2. Примерный механизм клеточной иммунной реакции
1. Чужеродные
антигены ГКГ-I на поверхности небольшого
количества клеток узнаются
Т-киллерами нескольких
антигенспецифических клонов. 2. Это вызывает активацию Т-киллеров
и их бласттрансформацию
3. Новые Т-киллеры атакуют
"чужеродные" клетки, выделяя при
этом белок перфорин.
4. В плазмолемме клеток образуются
гидрофильные поры, |
21.2.2.3. Примерный механизм гуморальной иммунной реакции
| 1. Небольшие количества
антигена (в свободном состоянии или в комплексе с
антителами, если таковые уже есть в организме)
поглощаются и перерабатываются антигенпредставляющими
клетками - В-лимфоцитами,
макрофагами или производными макрофагов. 2.
На поверхности этих клеток формируются
3. Стимулированные Т-хелперы, 4. Образуются плазмоциты 6. Связывание антител с антигенами может приводить:
|
21.2.3. Иммуноглобулины
а) А. Антигенная специфичность Т-клеток обусловлена наличием на их поверхности определённых антителоподобных (или Ig-подобных) рецепторных белков.
Б. Собственно же иммуноглобулины находятся на поверхности В-клеток, а также секретируются плазмоцитами.
б) Ниже
рассматривается структура иммуноглобулинов.
Что касается Ig-подобных рецепторов Т-клеток, они
во многом сходны по строению (хотя и имеют ряд
существенных особенностей).
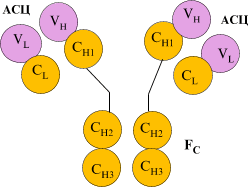
21.2.3.1. Принцип строения Ig
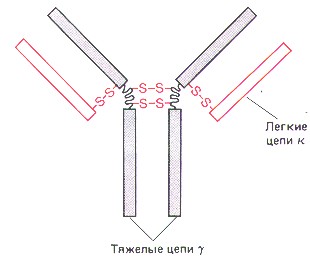
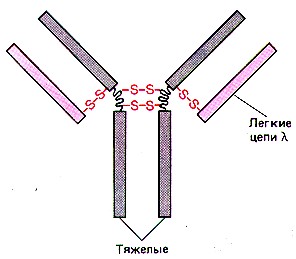
Общая структура антител. а) Строение антител отражается формулой: (L2H2)n ,
б) Каждая цепь включает несколько доменов (глобулярных участков): |
Схема - строение антител.
|
||||
а) Совокупность двух вариабельных доменов (VL и VН) составляет АСЦ - антигенсвязывающий центр. б) Как видно из схемы, в каждом антигене - не менее 2-х таких центров. в) Специфичность антитела (по отношению к тем или иным антигенным детерминантам) определяется именно вариабельными доменами, т.е. АСЦ. |
|||||
| Константные
участки. а) Постоянные (или константные) области на специфичность антигена не влияют. б) "Хвост" антитела, состоящий из четырёх константных доменов (СH2 и СH3 ), называется |
|||||
21.2.3.2. Классы иммуноглобулинов
| 1. Т.н. постоянные
области Н-цепей (включающие домены СH1, СH2 и СH3 ),
бывают 5 видов. 2. По
этому признаку все Ig (иммуноглобулины) подразделяются на 5 классов (обнаруживаемых в крови и различных
секретах организма):
|
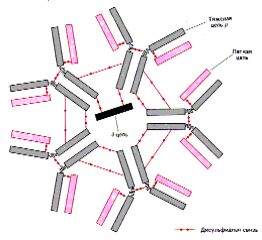
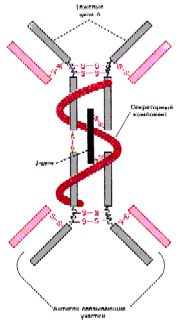
I. Иммуноглобулины класса M (Ig M)
| Особенности
структуры. 1. Ig M содержат тяжёлые цепи типа Нm. 2. Кроме того, эти Ig
3. Т.о., в 1 молекуле - 10 антигенсвязывающих центров. |
Структура: [L2 (Hm )2]5 J
|
| Образование IgM
до и после антигенной стимуляции. 1. а) Собственно, вначале клетки (нестимулированные В-лимфоциты) образуют мономерные молекулы Ig М - L2(Hm)2. б) Последние
2. И лишь после антигенной стимуляции образующиеся плазмоциты
начинают секретировать пентамерные молекулы Ig M. |
|
II. Иммуноглобулины класса G (Ig G)
| Особенности
структуры. Ig G - мономерные молекулы (n = 1) Образование. 1. Ig G образуются плазмоцитами
2. Поэтому Ig G - основной класс Ig в плазме крови (~ 70% от всех Ig плазмы). |
Структура: L2 (Hg )2
|
| СН-переключение. 1. Переход в течение первичного иммунного ответа с синтеза Ig M на синтез Ig G называется СН-переключением. 2. Его механизм тоже связан с перестройкой генома:
|
|
| Рецепторы к
FC-области. Как уже отмечалось,
благодаря чему комплексы IgG с антигенами (например, бактерии, покрытые IgG) связываются и фагоцитируются указанными клетками. |
|
III. Иммуноглобулины класса
А (Ig A)
| Локализация. 1. Ig А - основной класс Ig в секретах (молоке, слюне, слезах, секретах дыхательных путей и кишечного тракта). 2. Так, в п. 21.1.2.2 мы описывали появление плазмоцитов в слизистой оболочке кишечника. Исходная структура. Ig A, синтезируемые этими плазмоцитами, первоначально имеют структуру [L2 (Ha )2]2 J , т.е. являются димерами, содержащими (помимо лёгких цепей)
|
Структура:
|
| Связывание
секреторного компонента. 1. На базальной поверхности секреторных клеток имеется секреторный компонент S'.
2. Возможно, он защищает антитело от разрушительного действия ферментов, содержащихся в секретах. |
|
IV. Иммуноглобулины класса
Е (Ig Е)
| Структура. Ig Е - мономер с тяжёлыми цепями типа Нe .
Выше уже отмечалось, что
1. В норме содержание Ig E в плазме крови - очень низкое (меньше 0,01 % от всех Ig) . 2. При избыточной продукции Ig E наблюдаются аллергические реакции - бронхиальная астма, крапивница и т.д. |
Структура: L2 (He )2
|
V. Иммуноглобулины класса
D (Ig D)
| Структура: Ig D - мономеры с тяжёлыми цепями типа Нd .
1. Ig D
содержатся в
качестве рецепторов
(вместе с Ig M) на поверхности стимулированных В-клеток (плазмоцитов),
|
Структура: L2 (Hd )2
|
21.2.3.3. Система комплемента
| Функция и состав. 1. Если антитела
направлены против антигенов на поверхности циркулирующих в
крови бактерий, форменных элементов и т.д.,
2. В данной системе - около 20 белков плазмы крови, многие из которых -
|
|
Механизм действия.
Напомним: сходным образом уничтожают чужеродные клетки и клетки-киллеры (ТК , ТГЗТ , NK- и К-клетки; п. 20.2.5.4):
|
21.2.3.4. Первичный и вторичный ответ на антиген
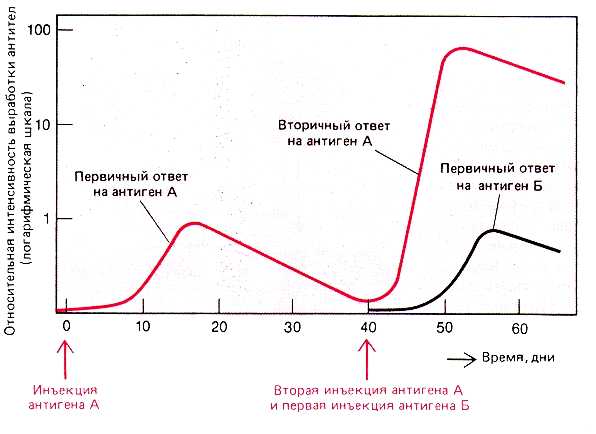
График - выработка антител (по оси ординат) при первичном и повторном введении антитела.
|
| Особенности
вторичного иммунного ответа. 1. При вторичной антигенной стимуляции (если она проводится не ранее, чем через 2-4 недели после первой) гуморальная реакция является гораздо более выраженной:
|
| Иммунологическая
память. 1. Таким образом, первичная стимуляция оставляет после себя иммунологическую память. 2. Механизмы этой памяти состоят в следующем:
|
![]()