

6.4. Функции нуклеиновых кислот
Функции ДНК определяются ее строением.
Основная функция ДНК заключается в хранении и передаче генетической информации. |
В молекулах ДНК с помощью особого кода зафиксированы все наследственные свойства организма. Участок ДНК, отвечающий за синтез одного белка (полипептида), назвали геном. Второе название гена - цистрон.
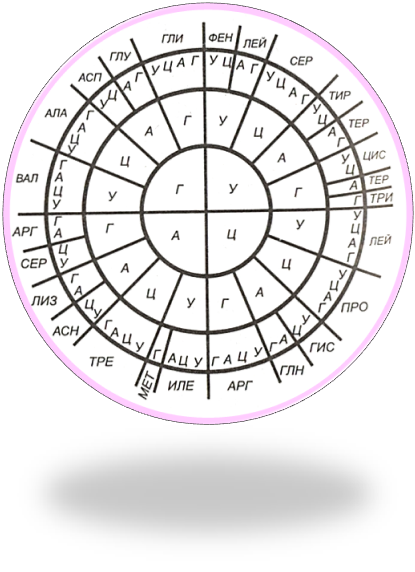
Уотсон и Крик максимально приблизили открытие генетического кода, установив структуру ДНК и механизм ее воспроизведения, но сделал это открытие другой ученый. В 1954 г. на суд биологов американский физик русского происхождения Георгий Гамов вынес гипотезу, которую можно назвать одной из самых революционных в биологии. Она провозглашала: информация, необходимая для синтеза белка, закодирована в генах. Порядок чередования 20 аминокислот в белке записан в молекуле ДНК кодом - чередованием четырех азотистых оснований: А, Г, Ц, Т. Гамов показал основные принципы кода. Три основания составляют триплет. Вырожденные триплеты кодируют одну и ту же аминокислоту (рис 6.3). Экспериментальное подтверждение теоретически рассчитанного генетического кода стало возможным лишь в 1961 г.
Рисунок 6.3. Генетический код Первая буква кодона расположена в центральном круге, вторая - в первом кольце и третья - во втором. В наружном конце записаны сокращенные названия аминокислот. |
|
Репликацию ДНК часто называют самоудвоением, хотя это и не совсем точно. На самом деле без ферментов, в т.ч. ДНК-полимеразы, ей не обойтись. Перед репликацией матричные цепи нужно разделить. Эту реакцию осуществляют два типа белков: хеликазы и SSB-белки (single-strand binding - белки, связывающиеся с однонитевой ДНК). Хеликазы - это АТФазы, использующие энергию гидролиза АТФ для расплетения двойной спирали (helix - спираль). Хеликаза “едет” по одной из цепей ДНК, расплетая перед собой двойную спираль. В результате работы хеликаз возникает репликационная “вилка” из двуспирального участка ДНК и двух одноцепочечных ветвей (рис. 6.4).
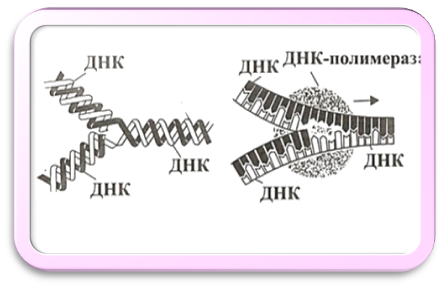
Рисунок 6.4. Репликационная вилка
Модель двойной спирали позволила понять принцип удвоения цепи ДНК, где каждая цепь служит матрицей, на которой выстраивается комплементарная ей (взаимодополняющая) новая цепь ДНК. Синтез ДНК на одноцепочечной матрице ведут ферменты ДНК-полимеразы. Ренатурации одноцепочечных участков ДНК препятствует их связывание SSB-белком, имеющим избирательное сродство к однонитевой ДНК. Связывание одноцепочечной ДНК с SSB-белком стимулирует ДНК-полимеразу и повышает точность ее работы.
На расплетенных участках синтез ДНК идет фрагментами. После синтеза фрагмента происходит дальнейшее расплетание цепей. Фрагменты сшиваются в одну непрерывную цепь с помощью еще одного фермента - ДНК-лигазы.
Исходя из изложенного, очевидны две основные функции ДНК:
1. Хранение генетической информации. Функция обеспечивается фактом существования генетического кода.
2. Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
Функции РНК. Известны три типа клеточной РНК. Первый тип - высокомолекулярная рибосомная РНК. Составляет около 85% клеточной РНК и отличается устойчивостью. Эта РНК выступает как структурный материал рибосом (на ее долю приходится 60, а на белок - 40%).
Второй тип РНК - информационная, переносящая генетическую информацию от ДНК хромосомы к месту синтеза белка, т.е. к рибосомам. Этот тип РНК и есть матрицы, с помощью которых “набираются” полипептидные цепочки белков. Поэтому ее также называют матричной РНК. В отличие от рибосомной она очень быстро синтезируется и распадается (от нескольких секунд до нескольких минут). В силу малого времени жизни ее концентрация в клетке составляет 3-5% всей клеточной РНК.
Третий тип РНК ѕ это низкомолекулярная транспортная РНК. Она специфически связывает и переносит аминокислоты и составляет около 12% всей рибонуклеиновой кислоты.
РНК, входящая в состав вирусов, выполняет четвертую функцию. Это геномная (вирусная) РНК, которая выполняет функцию хранения и передачи генетической информации вируса.
Таким образом, ДНК способна хранить генетическую информацию, удваиваясь при делении клетки, но не может передавать наследственную информацию к местам синтеза белка. Переносом информации из ядра в цитоплазму ведает и-РНК (информационная).
Информационная РНК рождается на ДНК по ее образу и подобию и несет на себе отпечаток генетического кода -- программу синтеза белка. Связываясь с рибосомами, и-РНК вносит в них эту программу и направляет действие рибосомы в сторону синтеза определенных белков. Комплементарные триплеты на и-РНК называются кодонами, а на транспортной РНК соответственно антикодонами.
Из клетки были выделены ДНК и и-РНК, и для доказательства их полной аналогии применили одновременное расплавление и последующее медленное охлаждение (отжиг) смеси ДНК и соответствующей и-РНК. Наблюдалось явление специфического связывания однотяжевых цепей ДНК и РНК. Интересно, что такую гибридную спираль ДНК - и-РНК не способны атаковать ни РНКаза, ни ДНКаза.
Ферментативный синтез и-РНК на матрице ДНК называется транскрипцией. |
Транскрипция - это первая стадия считывания генетической информации, на которой нуклеотидная последовательность ДНК копируется в виде нуклеотидной последовательности РНК. Транскрипция осуществляется ферментами РНК-полимеразами, синтезирующими РНК на ДНК-матрице. Следует отметить, что ДНК эукариот характеризуется прерывистой структурой генов. Участки ДНК, сохраняющиеся в зрелой РНК, называются экзонами. Экзоны разделяются интронами - участками ДНК, подлежащими выщеплению в процессе созревания. Только экзоны несут информацию, необходимую для синтеза белка. Функция интронов пока не ясна. А сам процесс созревания РНК из экзонов, исключающий интроны, называется сплайсингом (рис. 6.5).
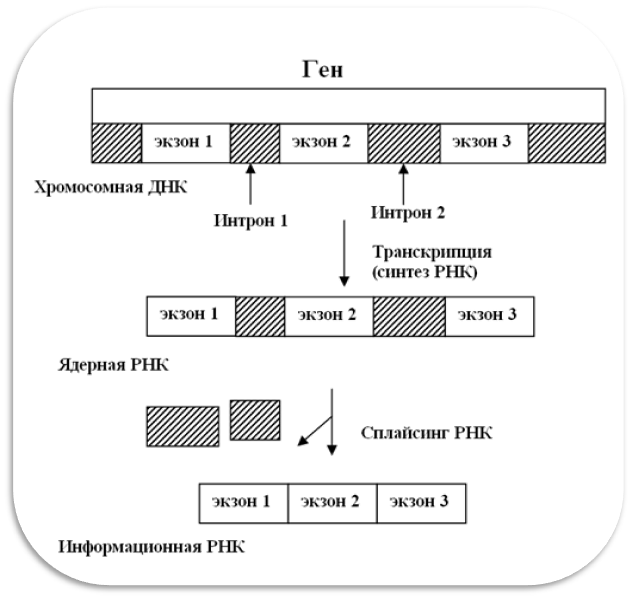
Рисунок 6.5. Сплайсинг
Синтез молекул РНК начинается в определенных местах ДНК, называемых промоторами, и завершается в терминаторах. Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции - оперон. Оказавшись на промоторе, РНК-полимераза образует с ним промоторный комплекс. Только с промотора может начаться синтез специфической РНК (рис 6.6). Диспетчерские функции выполняет оператор. Из нуклеозидтрифосфатов образуются первые несколько звеньев цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади - восстановление двойной цепи ДНК, одновременно нарастает цепь РНК.
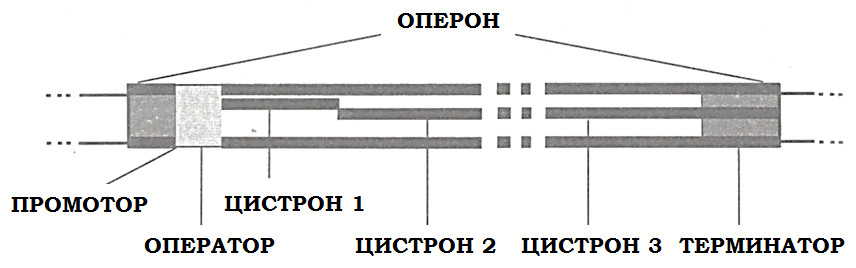
Рисунок 6.6. Строение оперона
В 1955 г. Ф. Крик предположил, что должны быть еще какие-то молекулы, заведующие транспортом аминокислот для их выстраивания в полипептидную цепочку. Это предположение подтвердилось экспериментально. Были открыты небольшие молекулы транспортных РНК. Для каждой аминокислоты есть своя т-РНК, доставляющая ее по месту назначения. Природа сделала так, что т-РНК «владеет двумя языками», одним концом она соединяется с аминокислотами, другим - с и-РНК. Молекула т-РНК подтаскивает одну аминокислоту к рибосоме и возвращается за другой.
Ферментативный синтез белка на матрице и-РНК называется трансляцией (рис 6.7). |
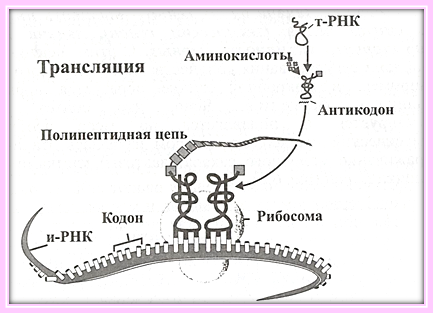
Рисунок 6.7. Биосинтез белка
В этом процессе главная функция транспортной РНК - связывание аминокислот в клетке для транспортировки их внутрь рибосом, где идет синтез белка. Каким образом, каждая молекула т-РНК узнает свою специфическую аминокислоту? Это узнавание осуществляется через посредство фермента (20 ферментов - по одному на каждую аминокислоту) и обозначают его словом рекогниция (recognition).
![]() Наконец, функции вирусных РНК. В чем они состоят?
Наконец, функции вирусных РНК. В чем они состоят?
Вирусы могут содержать как одноцепочечную РНК, так и двухцепочечную. Примером вирусов первого типа служат вирус гриппа, многие вирусы растений, второго - вирус цитоплазматического полиэдроза насекомых. Естественно, что функция вирусных РНК - это их репликация в организме хозяина. Когда РНК вируса однотяжевая, то, находясь в клетке хозяина, она способна действовать непосредственно как и-РНК, и хозяин начинает синтезировать белки вируса. Если РНК двухтяжевая, то сначала она расплетается и каждая цепь действует как и-РНК. В то же время существуют некоторые вирусы, которым для репликации необходимо сначала синтезировать ДНК на матрице РНК, а затем уже как обычно на ДНК - и-РНК. Для этих целей подобные вирусы имеют так называемую обратную транскриптазу - фермент, способствующий синтезу ДНК на матрице РНК.
Важнейшие функции обеих нуклеиновых кислот ДНК и РНК ярко проявляются при рассмотрении синтеза ферментов в клетке любого организма. Здесь мы подходим к понятию структурных и регуляторных генов.
Главные положения, лежащие в основе регуляции на генном уровне, были высказаны Ф. Жакобом и Ж. Моно. Суть гипотезы сводилась к тому, что не все гены хромосом одинаковы по своему значению. Задача одних генов - выдавать информацию на синтез белка (экспрессия генов), задача вторых - регулировать активность первых. С этим явлением связана проблема регулирования количества синтезируемого фермента. Например, в микробной клетке образуется порядка 1000 различных ферментов. Ясно, что синтез ферментов регулируется и подавляется по мере необходимости. Классический пример, на котором изучалась эта проблема - сбраживание молочного сахара (лактозы) клетками кишечной палочки E. coli (рис.6.8).
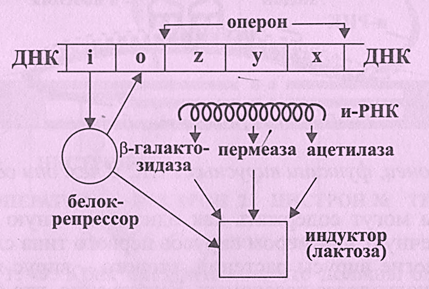
Рисунок 6.8. Регуляция биосинтеза белка:
z, y, x – структурные гены; i – регуляторный ген; o - оператор
Как только в клетке появляется субстрат - лактоза, она начинает вырабатывать фермент b-галактозидазу, расщепляющую лактозу на простые сахара, нужные клетке для питания. Эксперименты показали, что b-галактозидаза в клетке образуется всегда, но в очень малых количествах, и только при появлении лактозы синтез этого фермента сильно ускоряется, т.е. индуцируется. Поэтому лактозу можно назвать в данном случае индуктором. Отсюда следует также, что в хромосоме клетки E. coli имеется ген (обозначенный z), где закодирована информация о синтезе b-галактозидазы. Далее был найден дополнительный ген y (игрек), управляющий синтезом фермента пермеазы. Пермеаза осуществляет активный транспорт лактозы через клеточную оболочку. В присутствии пермеазы концентрация индуктора внутри клетки во много раз выше, чем вне ее. Оказалось, что оба гена z и y смежны и находятся рядом с третьим геном х (икс), образуя лактозный оперон, программирующий производство трех белков для расщепления лактозы. Оперону предшествует регуляторный участок ДНК - ген i, который синтезирует особый белок - репрессор. Репрессор имеет сродство и к генам структурным (составляющим оперон), и к гену-регулятору i, запирая то одни, то другие гены. Место действия репрессора на хромосому представляет собой небольшой участок в начале структурного гена z - оператор. Оператор выключает или включает секцию хромосомы, состоящую из генов z, y, x. Таким образом, белок-репрессор вхож и к оператору, и к индуктору (лактозе). Пока лактозы нет, репрессор блокирует оператор, и оперон выключен. Как только в клетке появляется индуктор, он соединяется с репрессором и лишает его возможности блокировать оператор, тогда оперон включается. Таким образом, мы имеем дело с саморегулированием: количество субстрата - входной сигнал, количество фермента - регулируемая величина. Отсюда можно сформулировать еще одну функцию нуклеиновых кислот как реализацию генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов.
Таким образом, достижения молекулярной биологии по изучению связей между структурой и функцией биологических полимеров - белков, ДНК и РНК - определили центральную догму ДНК↔РНК→белок. Функции нуклеиновых кислот и белков обеспечиваются процессами транскрипции и трансляции.
![]()
