Глава 3
· Внутриутробное развитие
Морфогенез — формирование пространственной организации организма и его частей. Многоклеточный организм развивается из оплодотворённой яйцеклетки (зигота) не только путём увеличения количества клеток (пролиферация) и увеличения массы зародыша (рост). Одновременно определяется судьба образующихся клеток, т.е. клетка «выбирает» один из многих возможных путей развития. Этот процесс известен как «детерминация». Детерминированные клетки специализируются (дифференцировка), т.е. приобретают определённую структуру и оказываются способными выполнять конкретную функцию. Одновременно осуществляется морфогенез — клетки координированным образом формируют органы и архитектуру пространственной организации тела. Формирование пространственной архитектуры зародыша (организма) и его частей (органов) осуществляется при реализации следующих морфогенетических процессов: направленная миграция клеток (в том числе направленный рост частей клеток, например, отростков нервных клеток), гибель клеток. Таким образом, пролиферация, рост, детерминация, дифференцировка, морфогенез, миграции клеток и их гибель — важные события (морфогенетические процессы) при развитии многоклеточного организма.
Причины развития
Всё многообразие клеток организма (разные типы клеток) развивается из одной единственной (зиготы). Говоря формально, все клетки организма — клон, родоначальником которого является оплодотворённая яйцевая клетка. Клон, по определению, совокупность идентичных клеток, развившихся из единственной клонообразующей клетки. Однако, для развивающегося организма это не так: существует множество разных клеточных типов, возникающих в результате различной — дифференциальной — активности генов. В чём же причина многообразия при исходной идентичности? В сущности, у нас один ответ — эндогенная программа многообразия в виде ядерного генома зиготы. Следовательно, генетическая программа — источник разнообразия клеток. Такое утверждение тривиально, но верно для одноклеточного зародыша. И это же утверждение становится неочевидным для многоклеточного зародыша, хотя бы в силу их различной взаимной позиции (например, на поверхности зародыша или в глубине, в левой или правой части зародыша, в верхней или нижней его части). Следовательно, для многоклеточного зародыша обязаны существовать внешние по отношению к конкретной клетке зародыша факторы, определяющие дифференциальную активность генов в разных клетках зародыша. Рассмотрение такой возможности определяет гипотеза позиционной информации в терминах «морфогенетическое поле», «левое–правое», «гомеобокс», «гомеозисные гены». Другими словами, возникновение разнообразия разных клеток в многоклеточном зародыше смещается в плоскость межклеточных взаимодействий («индукция», «индукционные взаимодействия», «факторы индукции»). Иными словами, дифференциальная активность генов определяется позицией конкретной клетки и реализуется при помощи разного типа межклеточных взаимодействий.
Детерминация
В эмбриогенезе появляются различия между клетками: возникают разные клеточные типы. Конкретные типы клеток образуют ткани. Из клеток разной тканевой принадлежности формируются органы. Определение пути развития тотипотентных клеток концептуса и полипотентных клеток концептуса, эмбриона, плода при внутриутробном развитии и организма в постнатальном онтогенезе происходит в ходе детерминации — процесса, в результате которого “компетентная клеточная система выбирает один из многих возможных путей развития” [61]. Такая потенциальная возможность развиваться в разных направлениях обозначается как проспективная потенция.
Дифференцировка
Дифференцировка — внешнее выражение детерминации. В ходе специализации конкретного клеточного типа (дифферон) формируются разные фенотипы клеток. Результат дифференцировки — специализированная клетка конкретной морфологии, выполняющая определённую функцию (состояние терминальной дифференцировки). По мере дифференцировки постепенно ограничиваются потенции клеток развиваться в различных направлениях. Дифференцировка необратима и осуществляется только в одном направлении — от менее дифференцированной к более дифференцированной структуре. При дифференцировке клетки экспрессируют строго определённую при детерминации часть генома: транскрибируют специфические РНК и синтезируют специфичные белки, что и определяет морфологические и функциональные признаки специализации клеток. Следовательно, различия между клетками, обладающими одинаковым набором генов, определяет дифференциальная активность генов.
Дифференциальная активность генов
Гены оплодотворённой яйцеклетки репрессированы. Начало развития сопровождается дерепрессией определённых групп генов (в первую очередь генов, контролирующих пролиферацию и общий метаболизм клетки). Первые тканеспецифические гены активируются на стадии гаструляции. Позднее, когда происходит координированная сборка сложных структур (органогенез), включаются другие гены. Разные клеточные типы экспрессируют разные гены.
Примеры переключения генов в ходе дифференцировки
Эритроидный клеточный тип. Гены глобинов: ранний эмбриональный (Hbf) ® эмбриональные Hb ® фетальный (HbF) ® два типа дефинитивных (HbА1 и HbА2).
Клетки хрусталика. Гены a-, b- и g-кристаллинов.
Миогенный клеточный тип. Гены миозина, тропомиозина, тропонина и других сократительных белков (разные для разных типов мышечной ткани).
Ограничение проспективных потенций. По мере дифференцировки постепенно ограничиваются потенции клеток развиваться в различных направлениях.
· Эритроидный клеточный тип. Зигота ® бластоциста ® внутренняя клеточная масса ® эпибласт ® мезодерма ® мезенхима ® стволовая кроветворная клетка (CFU-blast) ® полипотентная клетка–предшественница миелопоэза (CFU-GEMM) ® взрывообразующая единица эритропоэза (BFU-E) ® унипотентный предшественник эритроцитов (CFU-E) ® проэритробласт ® базофильный эритробласт ® полихроматофильный эритробласт ® нормобласт ® ретикулоцит ® эритроцит. Cтволовая кроветворная клетка (CFU-blast) — источник всех клеток крови. Полипотентная клетка–предшественница миелопоэза (CFU-GEMM) ограничена в потенциях и дифференцируется только в клетки миелоидного ряда. На следующем этапе происходит дальнейшее ограничение потенций. Так, взрывообразующая единица эритропоэза (BFU-E) — предшественница только одного типа клеток, а именно эритроидного.
· Клетки хрусталика. Зигота ® бластоциста ® внутренняя клеточная масса ® эпибласт ® эктодерма ® головная эктодерма ® хрусталиковая плакода ® хрусталиковый пузырёк ® хрусталик ® клетки хрусталика (синтез a- и b-кристаллинов) ® продвинутые клетки хрусталика (синтез a-, b- и g-кристаллинов). В ряду от зиготы до головной эктодермы происходит ограничение проспективных потенций клеток. Клетки головной эктодермы служат источником различных структур (например, хрусталиковой и слуховой плакод, зачатков лицевых структур). Потенции клеток хрусталиковой плакоды ограничены рамками одного дифферона. Они могут дифференцироваться только в клетки хрусталика.
· Миогенный клеточный тип. Зигота ® бластоциста ® внутренняя клеточная масса ® эпибласт ® мезодерма ® несегментированная мезодерма ® сомит ® клетки миотома ® миобласты митотические ® миобласты постмитотические ® мышечная трубочка ® мышечное волокно. В приведённой схеме от этапа к этапу ограничивается количество потенциальных направлений дифференцировки. Клетки несегментированной мезодермы имеют потенции к дифференцировке в различных направлениях и образованию миогенного, хондрогенного, остеогенного и других направлений дифференцировки. Клетки миотома сомитов детерминированы к развитию только в одном направлении, а именно к образованию миогенного клеточного типа (поперечнополосатая мышца скелетного типа).
Пролиферация и дифференцировка
Существенная черта дифференцировки состоит в том, что дифференцировка обычно наступает после пролиферации клеток. Быстро размножающиеся клетки, как правило, являются малодифференцированными (например, клетки базального слоя эпителия кожи или мезенхимные клетки). Наоборот, высокодифференцированные клетки, как правило, утрачивают способность к пролиферации (например, эритроциты и нейроны).
Примеры пролиферативной активности клеток по мере их дифференцировки
· Эритроидный клеточный тип. По мере дифференцировки, а её признаком служит способность синтезировать Hb, пролиферативная активность угасает. Проэритробласты не синтезируют Hb и многократно митотически делятся. Базофильный эритробласт активно синтезирует Hb и сохраняет способность к митозу. Полихроматофильный эритробласт содержит значительное количество Hb и также может делиться. Ранние нормобласты, заполненные Hb, по-видимому, ещё могут делиться, но постепенно утрачивают способность к делению и выталкивают ядро.
· Клетки хрусталика. В ходе закладки хрусталика предшественники дефинитивных хрусталиковых клеток активно делятся, выходят из клеточного цикла, синтезируют кристаллины и локализуются в центральной области хрусталикового эпителия. Увеличение количества клеток хрусталика происходит на протяжении всего онтогенеза за счёт герминативной зоны.
· Миогенный клеточный тип. Жёстко координированная последовательность сначала пролиферации, а затем дифференцировки наглядно проявляется для данного клеточного типа. Миобласты митотически делятся и только после этого переходят в постмитотические миобласты, останавливающиеся в фазе G0 и впредь не синтезирующие ДНК. Подобным митозам Хью Хольцер [94] дал название квантальные митозы. Они отличаются от обычных митозов (пролиферативные митозы) тем, что приводят к появлению новых морфогенетических свойств у клеточных потомков (в данном случае клетки приобретают способность к слиянию).
Необратимость дифференцировки
Дифференцировка необратима и осуществляется только в одном направлении — от менее дифференцированной к более дифференцированной структуре. Для нетрансформированных клеток, а также для стволовых клеток в условиях in vitro не следует применять термин дедифференцировка (как правило, речь при этом идёт о границах клеточного типа и его пластичности).
Морфогенетические процессы
Морфогенез осуществляется при реализации различных морфогенетических процессов: рост, межклеточные взаимодействия, индукция, направленная миграция клеток, направленный рост частей клеток (например, отростков нервных клеток), гибель клеток.
Рост
Рост — увеличение массы и, как правило, линейных размеров за счёт увеличения количества клеток, морфо-функциональных единиц органов, самих органов, систем органов и т.д. Увеличение массы без клеточных делений наблюдают при гипертрофии клеток в нормальных (например, гипертрофия хрящевых клеток, гипертрофия миометрия при беременности) и патологических условиях. В организме вырабатываются многочисленные гуморальные факторы, стимулирующие рост, а также пролиферацию различных клеточных типов, — факторы роста.
Межклеточные взаимодействия и индукция
Специализацию клеток и образование новых структур направляют межклеточные взаимодействия и индукция. Природу клеточных взаимодействий в морфогенезе объясняет концепция позиционной информации.
Позиционная информация
Общий план тела определяется очень рано. Позднее, на протяжении всего периода формирования органа или целого организма, детали морфогенеза уточняются при помощи сигналов позиционной информации. Согласно концепции позиционной информации, клетка «знает» своё местоположение в координатной системе зачатка органа и дифференцируется в соответствии с этим положением. Позиционную информацию клетка получает от других клеток. Более того, клетка достигает состояния терминальной дифференцировки только при условии своевременного получения ею серии последовательных сигналов позиционной информации. Зона, в пределах которой эффективно действуют сигналы позиционной информации, называется морфогенетическим полем. В течение ряда последующих клеточных делений клетки морфогенетического поля «помнят» о своем исходном назначении. Постоянная активность гомеозисных генов определяет в клетке память о позиционной информации.
В 1969 г. Льюис Вольперт [134] предложил модель позиционной информации («французского флага»), согласно которой положение клетки в морфогенетическом поле определяется в соответствии с существующей системой координат. Модель предполагает наличие специальных позиционных химических сигналов, или морфогенов, которые продуцируются клетками–источниками и формируют во внеклеточном пространстве диффузионные градиенты. Клетки–мишени регистрируют градиенты морфогенов и интерпретируют их для определения своей локализации в морфогенетическом поле. Существуют и другие представления о механизме, определяющем положение клеток в морфогенетическом поле. Так, согласно модели последовательной передачи сигнала от клетки к клетке (sequential cell context model), предполагается последовательная индукция различных сигналов в расположенных рядом группах клеток с последующим их совместным действием на экспрессию специфических генов в других прилагающих клетках.
Морфогены
Морфоген — сигнальная молекула, формирует паттерн в морфогенетическом поле, несёт позиционную информацию и оказывает влияние на клетки по концентрационному градиенту. Обычно это белок, который действует как транскрипционный фактор. К морфогенам относят многие секретируемые клетками белки, включая представителей различных семейств, в том числе Wnt и трансформирующего фактора роста b (TGFb). Название семейства Wnt происходит от англ. wingless — бескрылый и int — ген. Wnt был впервые открыт как протонкоген int1 (wingless and mouse mammary tumour virus integration site). Подобные белки связываются со своими рецепторами и активируют сигнальные каскады в клетках-мишенях, изменяя в них экспрессию специфических генов. Среди морфогенов, участвующих в индукции и спецификации, например мезодермы, упоминают активины, морфогенетические белки кости (BMP), факторы роста фибробластов (FGF). Градиенты морфогенов Sonic hedgehog (Shh) и Indian hedgehog (Ihh) продемонстрированы в ходе развития зубов, нервной трубки и сомитов. Представление о концентрационном градиенте морфогена предполагает, во-первых, прямое действие морфогена на клетку-мишень и, во-вторых, зависимость ответа данной клетки от концентрации морфогена.
Гомеозисные гены
Гомеозисные гены — семейство родственных генов, содержащих гомеобокс и определяющих форму тела. У млекопитающих это семейство представлено 38 генами, сгруппированными в 4 комплекса — Hox (Homeobox) A, Hox B, Hox C и Hox D (по номенклатуре Human Gene Mapping Workshops [рабочая группа картирования генов человека] соответственно Hox 1, Hox 2, Hox 3 и Hox 4). Существуют и другие комплексы, например, Hox 7. Эти группы генов расположены в хромосомах 2, 7, 12, 17. Гены экспрессируются в эмбриогенезе и определяют организацию общего плана тела. Экспрессия генов контролирует разделение тела эмбриона по координатным осям на морфогенетические поля. Транскрипты гомеозисных генов присутствуют в головном и спинном мозге, в почках конечностей и сердце с 5 по 9 неделю развития. Так, региональная специализация структур позвоночного столба направляется гомеозисными генами. Они контролируют пролиферацию и дифференцировку кроветворных клеток. Например, экспрессия генов комплекса Hox B (Hox 2) зарегистрирована в клетках эритромегакариоцитарных линий и популяций ранних предшественников гемопоэза.
· Гомеобокс — эволюционно консервативная последовательность, состоящая примерно из 180 пар нуклеотидов. Гены, содержащие гомеобокс, кодируют ядерные белки, регулирующие экспрессию генов, а гомеобокс кодирует часть ДНК–связывающего белка. Соответствующая последовательность из 60 аминокислотных остатков, кодируемая гомеобоксом, получила название гомеодомен. Гомеодомены входят в состав белков-регуляторов транскрипции, или гомеобелков. Гомеобелки содержат гомеодомен и связываются с ДНК, контролируя экспрессию генов. Эти важные транскрипционные факторы регулируют развитие эукариот. Гомеодомены расположены вблизи С–конца белка. Некоторые гомеобелки связываются с ДНК при помощи нескольких доменов. Структурные мотивы связывания с ДНК могут быть трёх разновидностей: "лейциновая застёжка", мотив "спираль-поворот-спираль" (helix-turn-helix, HTH) и «цинковые пальцы». Домен «лейциновая застёжка» содержит 4–5 остатков лейцина, разделенных семью другими аминокислотными остатками. Отличительной чертой HTH-мотива является то, что он размещается в большой бороздке двойной спирали ДНК и связывается с определённой нуклеотидной последовательностью. Фрагмент N–терминального конца гомеодомена располагается в малой бороздке ДНК. Белки «цинковых пальцев» играют важную роль в половой дифференцировке. Макромолекула может содержать более 10 повторов из 30 аминокислотных остатков, включающих 7–10 атомов цинка. Гомеобелки могут формировать многокомпонентные комплексы с другими транскрипционными факторами и проявлять свойства активаторов, эффекторов и репрессоров транскрипции. Известно до 25 классов гомеодоменов. В основе их классификации лежат два критерия: аминокислотная последовательность и доменный состав гомеобелков.
· Гомеозисные гены и врождённые аномалии. Впервые на связь гомеозисных генов и врождённых пороков указал Кэмпбелл (1989). Делеция короткого плеча хромосомы 4 как причина синдрома Вольфа-Хиршхорна сочетается с дефектами гена комплекса Hox 7. В частности, существует связь между дефектами гена этого комплекса и возникновением врождённой расщелины нёба.
Индукционные взаимодействия
В органогенезе — координированной сборке разных тканевых структур — важное значение имеют индукционные взаимодействия между эмбриональными зачатками. В ходе индукции клетки одного зачатка (источник) воздействуют на клетки другого зачатка (мишень). Источник инструктирует мишень к дифференцировке в конкретную структуру или разрешает дифференцировку. Возникшая структура оказывает индуцирующее влияние на другую мишень, и появляется новая структура и т.д. Эмбриогенез — сплошная череда индукционных взаимодействий.
· Первичная эмбриональная индукция — влияние хордомезодермы на дорсальную эктодерму; результат — образование зачатка нервной системы.
· Закладка конечностей. В результате индукционного воздействия клеток латеральной мезодермы на эктодерму возникает локальное утолщение эктодермы, вместе со скоплением мезодермальных клеток формирующее почку конечности.
· Формирование хрусталика. Воздействие выроста переднего мозга (глазного пузыря) на лежащую над ним эктодерму индуцирует образование в ней хрусталиковой плакоды, дающей начало хрусталику.
· Дифференцировка склеротома. Хорда и нервная трубка — источники индукционного влияния на клетки вентро-медиальной части сомита — склеротома. В результате клетки склеротома начинают интенсивно размножаться и покидают сомит, образуя зачатки позвонков, рёбер и лопаток.
Направленная миграция клеток
Зачастую эмбриональные клетки возникают и размножаются в одном месте, а дифференцируются и функционируют в другом. Поэтому из места своего образования клетки мигрируют в места окончательной локализации. Миграцию клеток в нужное место направляют сигналы микроокружения, специальные молекулярные метки. Носителями этих меток служат другие клетки или внеклеточный матрикс.
· Выселение клеток из гипобласта и образование внезародышевой энтодермы.
· Выселение клеток из нервного гребня с образованием меланоцитов, хромаффинных клеток, вегетативных и чувствительных нейронов и других структур.
· Выселение клеток из сомита. Клетки дерматома, наружного слоя дорсо-латеральной части сомита, выселяются под покровную эктодерму и формируют соединительнотканную часть кожи. Клетки склеротома также покидают сомит и образуют скелетные структуры.
· Направленная миграция клеток в нервной трубке. Специальные поддерживающие клетки радиальной глии возникают в раннем нейроонтогенезе и служат направляющими путями для адресной миграции нейробластов из внутреннего (эпендимного) слоя в строго определённые области наружных отделов нервной трубки. Так возникают различные слои нейронов в коре головного мозга.
Гибель клеток
Естественная (запрограммированная, физиологическая) гибель клеток — важный морфогенетический процесс на ранних этапах онтогенеза. Способ её реализации — апоптоз.
· Нейробласты. На строго определённых этапах развития нервной системы в зачатках различных её анатомических областей погибает от 25 до 75% общей численности популяции нейробластов.
· Развитие конечности. Межпальцевые участки почки конечности — зоны массовой гибели клеток. Синдактилия — дефект этого естественного процесса.
· Регрессия парамезонефрических протоков. В мужском организме под действием мюллерова ингибирующего фактора клетки парамезонефрических (мюллеровых) протоков (из них в женском организме развиваются маточные трубы, матка и часть влагалища) погибают, и протоки дегенерируют).
· Дегенерация пронефроса. Пронефрос — временная структура при образовании дефинитивной почки. Он индуцирует закладку мезонефроса, ещё одной промежуточной структуры при закладке почки. Выполнив свою задачу, пронефрос дегенерирует.
Молекулярные механизмы морфогенеза
Дифференциальная активность генов и избирательные взаимодействия клеток являются основными концептуальными понятиями, объясняющими механизмы индивидуального развития. В основе дифференциальной активности генов лежат обратимые (эпигеномные) изменения отдельных участков хроматина с сохранением непрерывной структуры генома (конденсация, деконденсация хроматина, метилирование ДНК), а также необратимые изменения участков хроматина, которые могут быть связаны с частичной перестройкой генома. Экспрессия генов регулируется на разных уровнях: транскрипции, посттранскрипционном, трансляции мРНК, посттрансляционном созревании белков.
Дифференцировка клеток и морфогенез осуществляются в ходе избирательного взаимодействия клеток, которое реализуется с участием диффузных молекулярных сигналов. В подобных рецептор-опосредованных взаимодействиях клеток участвуют паракринные (вырабатываемые в непосредственной близости) факторы, факторы роста, многочисленные низкомолекулярные агенты. При избирательном взаимодействии клеток передача сигналов осуществляется не только при помощи паракринных сигналов, но и путём поверхностных белок–белковых взаимодействий, ферментативных реакций постсинтетической модификации белков (фосфорилирования, специфического расщепления и др.).
Список биологически активных макромолекул, участвующих в реализации генетической программы молекулярной спецификации клеток, достаточно велик. Подобные молекулярные сигналы сгруппированы в семейства (табл. 3-1). Рассмотрим некоторые из них, участвующие в ключевых морфогенетических процессах.
Надсемейство трансформирующего фактора роста b
Трансформирующий фактор роста b (TGFb)— белковый фактор роста, действующий на клетки-мишени через рецепторы, встроенные в плазмолемму. TGFb служит сигналом, контролирующим пролиферацию и дифференцировку клеток многих типов. Надсемейство TGFb включает большое количество различных белков, которые взаимодействуют с другими факторами роста и стимулируют или угнетают их активность. К надсемейству TGFb относятся морфогенетические белки кости (BMP), которые впервые были описаны как индукторы формирования кости и хряща. BMP участвуют в дорсо-вентральной организации мезодермы. BMP4 индуцирует образование вентральной мезодермы, включая кровь.
Семейство фактора роста фибробластов
Фактор роста фибробластов (FGF) содержит не менее 17 секретируемых клетками белков. Эти лиганды взаимодействуют с рецепторами, связанными с тирозинкиназой. Участвуют в спецификации клеток различных тканей. FGF был первым очищенным фактором, для которого была показана способность индуцировать образование мезодермы. FGF необходим для формирования туловищной и задней мезодермы, включая нотохорд и мышцы. Рецепторы FGF объединены в семейство белков, объединяющее 4-х гомологичных представителя. Молекула рецептора FGF состоит из 3-х фрагментов. Внеклеточный фрагмент представлен тремя иммуноглобулин-подобными доменами. Он связан с внутримембранным фрагментом, к которому со стороны цитоплазмы примыкает тирозинкиназный фрагмент молекулы. Во взаимодействии лиганда с рецептором участвует большая и разветвлённая молекула гепарансульфат протеогликана (рис. 3-1).
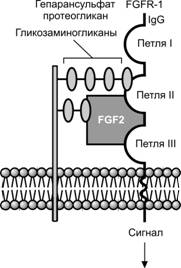
Рис. 3-1. Взаимодействие фактора роста фибробластов 2 (FGF2) с рецептором. FGF2 связывается со своим рецептором, встроенным в клеточную мембрану, при участии специального молекулярного посредника — гепарансульфат протеогликана, без которого взаимодействие лиганда с рецептором оказывается невозможным. Разветвленная молекула гепарансульфат протеогликана «прижимает» лиганд (FGF2) к иммуноглобулин-подобным доменам в наружной части молекулы рецептора фактора роста фибробластов 1 (FGFR1). Молекула FGF2 одной своей поверхностью связана с молекулой рецептора, а другой взаимодействует с ветвью гликозаминогликана, принадлежащего макромолекуле гепарансульфат протеогликана. Другая более длинная ветвь гликозаминогликана непосредственно «причалена» к молекуле рецептора и удерживает её, а белковая часть протеогликана интегрирована в клеточную мембрану. [65]
Wnt-родственные молекулы
Семейство включает гены, родственные гену wingless сегментарной полярности у Drosophila. Продукты этих генов, Wnt–белки, являются ключевыми регуляторами эмбрионального развития. Многие рецепторы и ингибиторы вмешиваются в Wnt–сигнальный путь, регулируя в конечном итоге стабильность белка b–катенина. Этот сигнальный каскад контролирует поддержание популяции стволовых клеток, процессы адгезии клеток и их миграции, раннего эмбриогенеза, гистогенеза, формирования суставов в ходе развития конечностей и связан с возникновением опухолей. У человека известно 4 гена Wnt. Ген Wnt7a кодирует синтез секреторного гликопротеина 349aa. Из рецепторов Wnt наиболее изучены представители семейства, кодируемые геном Frizzled. Семейство образовано семью различными трансмембранными рецепторами.
Семейство Hedgehog
У позвоночных представлено тремя белками: Sonic hedgehog (Shh), Indian hedgehog (Ihh) и Desert hedgehog (Dhh). Белки Hedgehog модифицируются в ходе аутокаталитического процессинга и становятся активными сигналами. С N–концевым доменом белка связывают все известные сигнальные свойства молекулы, тогда как C–концевой домен выступает в роли холестерол трансферазы. Hedgehog действует через рецепторы двух типов: Patched (Ptc) (12 трансмембранных доменов) и Smoothened (Smo) (7 трансмембранных доменов). Hedgehog вовлечён в морфогенез различных частей тела. Он участвует в нейрогенезе, гемопоэзах, закладке кости, формировании гонад и других структур.
· Sonic hedgehog (Shh) является фактором роста. Ген Sonic hedgehog гомологичен гену Hedgehog, который у дрозофилы выполняет функции гена сегментарной полярности. Этот ген был впервые описан у мутантов дрозофилы с проявлением выраженных изменений в областях, которые имеют отношение к формированию обнажённой поверхности кутикулы. Shh связывается с мембранным рецептором Patched (Ptc). Действие Shh зависит от места — источника сигнала, расстояния от клетки-источника до клетки-мишени и от возможного протеолитического его расщепления. В ходе развития конечности Shh выделяется клетками зоны поляризующей активности, которая контролирует формирование осей. Shh участвует в детерминации дорсо-вентральной полярности нервной трубки, контролируя развитие вентральной её части. Shh участвует в закладке некоторых отделов головного мозга, средней части лица, волосяных фолликулов. Shh определяет симметрию тела «правое-левое», детерминирует передне-заднюю ось зачатка конечности. Shh участвует в региональной спецификации кишки и контролирует форму кончика зуба.
à Рецепторы Sonic hedgehog. В ответ на действие сигнала Shh рецепторы Patched (Ptc) и Smoothened (Smo) объединяются. Рецептор Patched (Ptc) сдерживает развитие опухолей, т.к. инактивация этого рецептора приводит к их появлению. В половозрелом организме рецептор Patched (Ptc) экспрессируется в мозге, лёгких, печени, сердце, плаценте, скелетной мышце, поджелудочной железе и почке. В ходе развития рецептор Patched (Ptc) вместе с Shh присутствует в вентральной части нервной трубки, сомитах, в пределах зоны поляризующей активности развивающейся конечности. Дефекты Patched (Ptc) приводят к развитию синдрома базальноклеточного невуса (синдрома Горлина) — множественной карциномы кожи с кистами челюстей, эритематозными углублениями на ладонях и стопах и аномалиями скелета, особенно лицевого. Дефект рецептора Smoothened (Smo) может быть причиной наиболее распространенной базальноклеточной карциномы.
à Дефект гена Shh является причиной голопрозэнцефалии (тип 3), наpушения диффеpенциpовки переднего мозга на полушаpия и доли. Эта аномалия может сочетаться с анофтальмией (циклопией), недоразвитием носа и верхней губы, отсутствием обонятельного нерва и мозолистого тела.
· Indian hedgehog (Ihh) выполняет по крайней мере две функции.
à Первая функция связана с контролем развития энтодермы. На ранних стадиях закладки этого зародышевого листка Ihh определяет формирование висцеральной его части. На поздних стадиях Ihh регулирует пролиферацию и дифференцировку стволовых клеток в эпителии слизистой оболочки кишки.
à Другая функция Ihh связана со стимуляцией формирования кости в постнатальном периоде, когда кости удлиняются особенно быстро. К концу первого года жизни кости удлиняются на 50%, а к концу третьего года только на 7%. Ihh, действуя вместе с пептидом, связанным с паратиреоидным гормоном (PTHrP), участвует в разблокировании механизмов дифференцировки хондроцитов. Хондроциты между стадиями пролиферации и гипертрофии начинают секретировать Ihh и экспрессируют рецепторы для пептида, связанного с паратиреоидным гормоном (PTHrP). Ihh поддерживает экспрессию белков Gli и Patched (Ptc) в надкостнице, прилежащей к зоне гипертрофии хряща. Эти белки индуцируют синтез пептида, связанного с паратиреоидным гормоном (PTHrP), который, в свою очередь, сдерживает рекрутирование новых хондроцитов для хондрогенной дифференцировки. Делеция гена Ihh приводит к ахондроплазии — системному поражению скелета — врождённой болезни, характеризующейся нарушением энхондрального остеогенеза, карликовостью, короткими конечностями при обычной длине туловища вследствие угнетения пролиферации хондроцитов и их интенсивной гипертрофии. Таким образом, Ihh рассматривают как главный координатор роста кости и формирования костного скелета.
à Наконец, Ihh, действуя через мембранные рецепторы Ptc и Smo, стимулирует экспрессию фактора транскрипции Tal-1 и рецепторов VEGF (Flk1 и Flt1). Эти сигнальные пути прослежены в ходе дифференцировки гемангиобластов в желточном мешке (см. ниже).
· Desert hedgehog (Dhh) участвует в регуляции ранних и поздних событий сперматогенеза и дифференцировке глиальных клеток. Продукт гена Dhh появляется в предшественниках клеток Сертоли тотчас перед экспрессией ими детерминирующего фактора семенника Sry. В нервной системе Dhh вырабатывается шванновскими клетками. Секретируемая молекула действует на расположенные рядом фибробласты через рецепторы Patched (ptc), активируя процесс формирования соединительнотканного каркаса периферического нерва.
Сигнальный путь DSL/Notch
Гены семейства Notch кодируют структурно консервативные последовательности для поверхностных мембранных рецепторов, которые связываются с лигандами из семейства DSL (Delta/Serrate/Lag2). Этот сигнальный путь участвует в спецификации клеток и установлении границ между группами клеток различного типа. Этот путь значим для нейрогенеза, гемопоэзов, включая дифференцировку T–лимфоцитов, миогенеза, ангиогенеза, закладки зубов. Дефекты этого сигнальном пути приводят к возникновению опухолей. Notch белки являются трансмембранными рецепторами для трансмембранных лигандов Jagged и Delta. Во внеклеточных доменах белков Notch, Delta и Jagged присутствуют повторы белка эпидермального фактора роста (EGF). Notch действует, с одной стороны, как лиганд-связывающий рецептор, а с другой стороны, как транскрипционный фактор. Когда лиганд связывается с внеклеточным доменом, молекула вступает в цикл последовательных расщеплений, в результате которых цитоплазматический домен транслоцируется в ядро, где активирует Notch-зависимые гены.
Дефекты Notch, Jagged и Delta
· Мутации гена Notch3 вызывают синдром CADASIL (Cerebral Autosomal Dominant Arteriopathy with Subcortical Infarcts and Leukoencephalopathy), который проявляется нарушением мозгового кровообращения, деменцией, судорогами, депрессией, нарушением двигательных функций, параличом и инфарктами белого вещества мозга. При мутациях и транслокациях генов Notch2 и Notch3 велик риск развития злокачественных новообразований.
· Jagged-1 служит лигандом Notch–белков. Мутации гена Jag1 являются причиной синдрома Аладжила (множественные аномалии развития печени, выступающий лоб, заострённый подбородок, дефекты пигментации сетчатки, клапанов сердца, формы позвонков, задержка физического, умственного и полового развития).
· Мутации гена Jag2 — причина перинатальной смертности вследствие дефектов морфогенеза черепно-лицевой области в виде расщелин нёба, срастания языка с нёбными занавесками и сочетаются с синдактилией и дефектами развития тимуса.
· Мутации гена delta-подобного лиганда-3 приводят к спондило-рёберному дизостозу с характерными аномалиями позвонков и деформацией рёбер, затруднением дыхания и высокой смертности вследствие респираторной инфекции.
Другие морфогены и молекулы, контролирующие паттерны и судьбу клеток
Ноггин (Noggin). Секреторный белок, экспрессируется на разных этапах развития, начиная с зиготы и выполняя на каждом конкретном этапе определенную задачу. Важен для морфогенеза хряща и формирования суставов. Ингибитор сигнальных путей BMP, регулирующих развитие нервной трубки и сомитов. Дефекты ноггина являются причиной множественных синостозов (сращений костей), анкилоза (тугоподвижность сустава) стремечка в сочетании с увеличением больших пальцев верхних и нижних конечностей, проксимального симфалангизма, характеризующегося отсутствием проксимальных межфаланговых суставов. Нарушение экспрессии гена ноггина приводит к дефекту слуха вследствие срастания стремечка с каменистой частью височной кости.
Гремлин-1 (Gremlin-1). Цитокин, играет важную роль в канцерогенезе и развитии окончательной почки (метанефроса). Являясь антагонистом BMP, участвует в закладке конечностей на ранних этапах, взаимодействует с сигнальными каскадами FGF4–Shh. Дозозависимым путём угнетает сигнальный путь BMP4. Ингибирует хемотаксис моноцитов.
Таблица 3-1. Наиболее значимые гены и кодируемые ими сигнальные молекулы, участвующие в морфогенетических процессах
|
Гены\морфогены и их характеристика |
|
Гомеозисные гены (Hox) образуют 4 генных кластера (Hox A, B, C, D), которые организованы в 13 гомологичных групп (паралогов) |
|
Wnt. Wnt белки являются ключевыми регуляторами эмбрионального развития, контролируют поддержание популяции стволовых клеток, процессы адгезии клеток и их миграции, раннего эмбриогенеза, гистогенеза, формирования суставов в ходе развития конечностей и связан с возникновением опухолей |
|
Sonic hedgehog (Shh) контролирует развитие вентральной части нервной трубки, некоторых отделов головного мозга, средней части лица, конечностей, волосяных фолликулов, зубов. Дефект гена Shh приводит к нарушению формирования переднего мозга — голопрозэнцефалии (тип 3) |
|
Paired Box (Paired axial) (Pax). Название гена и его продукта связано с наличием парного домена (paired box sequence) в составе молекулы белка. Гены принадлежат семейству Homeobox и кодируют транскрипционные факторы Pax, которые гомологичны продуктам генов сегментации paired и gooseberry дрозофилы. У млекопитающих насчитывают до 9 белков Pax. Ген Pax3 экспрессируется в нервной трубке, нервном гребне и в дермамиотоме сомита. Pax3 контролирует закрытие нейропоров, ингибируя p53-зависимый апоптоз. Мутации Pax3 приводят к дефектам нервной трубки, а также к возникновению опухолей. Кроме того, они рассматриваются как причина синдрома Варденбурга (Waardenburg) — группы наследственных множественных врождённых пороков развития с характерным нарушением пигментации и глухотой. Pax1 контролирует процесс сегментации и формирование осевых структур. Мутации Pax1 приводят к аномалиям позвоночного столба по всей его длине. Pax2 контролирует дифференцировку клеток почки. Pax5 служит транскрипционным фактором B–лимфоцитов и регулирует в них экспрессию CD19. Pax5 участвует в нейрогенезе и сперматогенезе. Pax6 (окулоромбин) контролирует развитие структур глаза и носа. Нарушение экспрессии Pax6 является причиной аниридии II типа, полное или частичное отсутствие радужки |
|
Фактор роста фибробластов (FGF). Семейство содержит не менее 17 секретируемых клетками белков. Эти лиганды взаимодействуют с рецепторами, связанными с тирозинкиназой. Участвуют в спецификации клеток различных тканей. FGF участвуют в формировании туловищной и задней мезодермы, включая нотохорд и мышцы |
|
Трансформирующий фактор роста b (TGFb). Трансформирующий фактор роста b — белковый фактор роста, действующий на клетки-мишени через рецепторы (серин/треониновые киназы), встроенные в плазмолемму. TGFb служит сигналом, контролирующим пролиферацию и дифференцировку клеток многих типов. Надсемейство TGFb включает большое количество различных белков, которые взаимодействуют с другими факторами роста и стимулируют или угнетают их активность |
|
Морфогенетический белок кости (BMP) принадлежит надсемейству TGFb. Индуцирует образование хряща и кости. Участвует в развитии эндокарда (эндокардиальных подушек) |
|
Сосудистый эндотелиальный фактор роста (VEGF) принадлежит семейству фактора роста из тромбоцитов (PDGF), секретируется, но остается связанным с поверхностью клеток или внеклеточным матриксом. Один ген кодирует синтез четырёх молекулярных форм, которые образуются в процессе альтернативного сплайсинга |
|
Ретиноевая кислота — мощный морфоген, взаимодействует с цитоплазматическими рецепторами, после чего транслоцируется в ядро, где связывается с ДНК. В рецепторе ретиноевой кислоты различают три домена: N–концевой, ДНК–связывающий и C–концевой домен, взаимодействующий с лигандом |
|
Engrailed (En). Гены кодируют транскрипционные факторы, действие которых избирательно проявляется в зачатке конечности эмбриона в виде активации или ингибирования других генов. Принадлежат семейству Homeobox |
|
SRY — белок детерминации мужского пола. Ген характеризуется очень низкой дозой |
|
Гены T-Box (Tbx). Семейство транскрипционных факторов с избирательной экспрессией в ходе закладки верхних и нижних конечностей. Tbx5 участвует в развитии сердца |
Периоды пренатального развития
В пренатальном развитии человека выделяют начальный, зародышевый и плодный периоды (табл. 3-2).
Таблица 3-2. Периоды пренатального развития
|
Период |
Продолжительность периода |
Критическое событие (начало периода) |
Другие важные события периода |
|
Начальный (концептус) |
Первые 2 недели |
Оплодотворение |
|
|
Зародышевый (эмбрион) |
3–8 недели |
Образование первичной полоски |
Органогенез |
|
Плодный (плод) |
До конца беременности |
Плацентация |
|
* Считают, что эмбрион — общность клеток или существо, формирующееся на стадии первичной полоски, но не ранее. По мнению A. McLaren [69], нельзя называть эмбрионом продукт дифференцировки зиготы до стадии первичной полоски. Автор предлагает называть этот продукт концептусом, проэмбрионом, предэмбрионом.
Для начального периода и раннего эмбриона (см. рис. 3-5) выделены такие фазы развития и связанные с ними события: зигота — начало синтеза ДНК и белка; дробление — начало синтеза основных типов РНК; морула — клетки ещё тотипотентны (эмбриональная стволовая клетка); бластоциста — утрата тотипотентности; клетки детерминированы к образованию зародышевых или внезародышевых структур; гаструла — наличие зародышевых листков и стволовых клеток; органогенез — формирование зачатков органов из клеточных клонов.
Оплодотворение — слияние мужской и женской гамет, приводящее к образованию одноклеточного зародыша — зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы; при этом сливаются их ядра (пронуклеусы), объединяются хромосомы, и возникает диплоидная клетка нового организма — зигота. Начало оплодотворения — момент слияния мембран сперматозоида и яйцевой клетки, окончание оплодотворения — момент объединения материала мужского и женского пронуклеусов. Всё, что происходит до слияния мембран сперматозоида и яйцевой клетки, именуют событиями, предшествующими оплодотворению.
В эякуляте человека содержится 3´108 сперматозоидов (рис. 3-2). В женских половых путях они сохраняют способность к оплодотворению максимально до 2 суток. Примерно 200 из них достигает воронки маточной трубы, где происходит встреча сперматозоида с яйцеклеткой (см. позицию 2 на рис. 3-6).
Акросома образуется в ходе сперматогенеза как продукт комплекса Гольджи и может рассматриваться как аналог лизосом. Акросома (рис. 3-2) расположена в головке сперматозоида, кпереди от ядра и тотчас под плазматической мембраной. Спереди мембрана акросомы (наружная) соприкасается с клеточной мембраной сперматозоида, а сзади (внутренняя мембрана) — с ядерной мембраной.
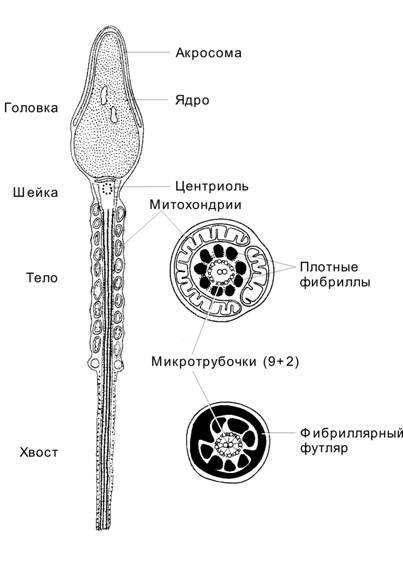
Рис. 3-2. Сперматозоид. Головка содержит ядро и акросому. В шейке присутствует центриоль. Промежуточный отдел заполнен митохондриями, формирующими митохондриальную спираль. В осевой части промежуточного отдела и практически вдоль всего хвоста располагается аксонема. В хвосте (снаружи от аксонемы) локализуются наружные плотные фибриллы и фибриллярный футляр. [17]
Направленная миграция
В феномене встречи сперматозоида и яйцеклетки важную роль играет хемотаксис — направленная миграция сперматозоидов по градиенту химических веществ, выделяемых овулировавшей яйцеклеткой. Хемоаттрактанты попадают в маточную трубу в составе фолликулярной жидкости лопнувшего фолликула, а также секретируются яйцеклеткой и фолликулярными клетками corona radiata.
· Обонятельне рецепторы hOR17-4, локализованные в мембране хвоста сперматозоида, отвечают за хемотаксис. Связанные с G–белком (Gaolf) рецепторы hOR17-4 активируют аденилатциклазу, запускающую внутриклеточный каскад на повышение содержания Ca2+ в цитоплазме хвоста. Повышение концентрации Ca2+ модулирует движение хвоста сперматозоида.
· Пахучее вещество bourgeonal, имеющее запах цветка лилии, специфически связывается с рецептором hOR17-4 и является сильным хемоаттрактантом для сперматозоидов. На основе химической формулы bourgeonal ведутся поиски естественных хемоаттрактантов сперматозоидов.
· Прогестерон считается слабым хемоаттрактантом, но за счёт медленного повышения концентрации ионов кальция в теле сперматозоида гормон повышает двигательную активность клетки.
Капацитация
До того, как сперматозоид встретится с яйцеклеткой, он в течение нескольких часов находится в женских половых путях. При этом на сперматозоид воздействуют факторы женского организма (pH, слизь, прогестерон, хемоаттрактанты и др.), поддерживающие способность к миграции и оплодотворению. Жизнеспособные подвижные сперматозоиды не способны оплодотворить яйцеклетку пока не пройдут окончательное созревание в женских половых путях. Процесс, при котором сперматозоид приобретает способность к оплодотворению яйцеклетки называется капацитацией. Только после капацитации сперматозоиды смогут связаться с прозрачной оболочкой, осуществить акросомную реакцию, проникнуть в яйцеклетку и оплодотворить её. Для успешной капацитации сперматозоиды, должны примерно 7 часов находиться в женских половых путях.
Механизм капацитации. Капацитация представляет собой сложный молекулярный механизм, подготавливающий мембрану сперматозоида для связывания с прозрачной оболочкой, акросомной реакции и оплодотворения. При капацитации усиливается фосфорилирование остатков тирозина мембранных и цитозольных белков сперматозоида, благодаря чему повышается его двигательная активность и готовность к акросомной реакции. Лишь небольшая популяция сперматозоидов одновременно вступает в капацитацию, которая длится в течение 1–4-х часов. Поэтому в состоянии ожидания яйцеклетки на смену первой популяции сперматозоидов приходит другая и т.д., для того чтобы в любой момент быть готовым для оплодотворения.
· Прогестерон и фолликулярная жидкость инициируют два основных события, стимулирующих фосфорилирование остатков тирозина. Это снижение содержание холестерина (и других стеринов) в плазматической мембране сперматозоида и образование активных форм кислорода. Супероксид анион O2– регулирует цАМФ-зависимое фосфорилирование остатков тирозина. Выход холестерина из мембраны увеличивает её текучесть и проницаемость, что облегчает фосфорилирование тирозина в мембранных белках.
· Последовательный ход молекулярных событий можно представить следующим образом: прогестерон (фолликулярная жидкость) ® образование супероксид аниона (O2–) и выход из мембраны холестерина ® активация аденилатциклазы ® увеличение содержания цАМФ ® активация протеинкиназы А (PKA) ® фосфорилирование остатков тирозина ® капацитация.
Механизмы предотвращения преждевременной капацитации
· Семеногелины I и II ¾ основные белки семенной жидкости, продуцируются в семенных пузырьках, образуют гелеобразный матрикс, заключающий в себя сперматозоиды, ферменты, фруктозу, витамины, металлы и др. Белки расщепляются с помощью простато-специфического антигена (PSA) с образованием фрагментов, вследствие чего, с одной стороны, происходит разжижение спермы и обеспечивается подвижность сперматозоидов, а с другой ¾ предотвращается капацитация незрелых сперматозоидов путём связывания активных форм кислорода.
· Гликоделины-S, -A и -F ¾ семейство гликопротеинов, ингибирующих капацитацию. Гликоделин-S содержится в семенной жидкости, блокирует капацитацию сперматозоидов до эякуляции. Белок специфически связывается с головкой сперматозоида и снижает утечку холестерина из мембраны. Гликоделин-А секретируется клетками эндометрия и маточных труб, блокирует связывание сперматозоида с прозрачной оболочкой яйцеклетки, а также защищает сперматозоиды от агрессии со стороны иммунной системы в женских половых путях. Гликоделин-F содержится в фолликулярной жидкости, ингибирует прикрепление сперматозоидов к прозрачной оболочке, подавляет индуцированную прогестероном акросомную реакцию.
Вызванная капацитация является одним из условий для успешного оплодотворения in vitro. В настоящее время предлагаются коммерческие среды, стимулирующие капацитацию.
Яйцеклетка
Большинство яйцеклеток в яичниках находится на стадии поздней диплотены профазы первого деления мейоза. При разрыве зрелого фолликула (овуляция) из яичника в брюшную полость выбрасывается овоцит второго порядка (рис. 3-3). Его окружают прозрачная оболочка (zona pellucida) и лучистый венец (corona radiata). Между прозрачной оболочкой и плазмолеммой яйцеклетки находится первое полярное тельце.
Перемещение яйцеклетки
Перед овуляцией увеличение тонуса ГМК маточной трубы приближает воронку маточной трубы к поверхности яичника. Биение ресничек эпителиальных клеток фимбрий ампулы маточной трубы создаёт ток жидкости, способствующий перемещению яйцеклетки из брюшной полости в ампулу.
При синдроме Картагенера и синдроме неподвижных ресничек женщины фертильны.
Способность к оплодотворению. Для успешного оплодотворения яйцеклетка должна встретиться со сперматозоидом в течение одних суток после овуляции. Из практических соображений время, в течение которого овулировавшая яйцевая клетка может быть оплодотворена, оценивают в пять суток. В течение этого времени яйцеклетка находится в состоянии пониженной активности и если оплодотворение не наступит, она погибает.
Прозрачная оболочка
По мере развития яйцеклетки в ней происходят синтез и последующая секреция гликопротеинов, постепенно формирующих zona pellucida. Зрелая прозрачная оболочка содержит густую сеть тонких нитей, состоящую из гликопротеинов (главным образом, белки ZP); один из них (ZP3) — главный рецептор сперматозоида. Инактивация или отщепление О-олигосахарида от молекулы ZP3 блокирует связывание сперматозоида с яйцеклеткой. ZP2 — вторичный рецептор сперматозоида, обеспечивающий дополнительное связывание гамет. Связывание сперматозоида с гликопротеинами ZP является сигналом к акросомной реакции.
Zona pellucida подразделяется на два слоя: внутренний, богатый нейтральными гликозаминогликанами, и наружный, содержащий преимущественно кислые гликозаминогликаны. В состав прозрачной оболочки также входят сульфатированные гликозаминогликаны, гиалуроновая и сиаловые кислоты, гликопротеины. Основная масса зрелой прозрачной оболочки — гликопротеины ZP: ZP1, ZP2 и ZP3 с Mr 90–110, 64–76 и 57–73 кД соответственно. ZP3 состоит из полипептида (44 кД), цепей N–олигосахаридов и О-олигосахаридов. Синтез ZP прекращается в ходе или тотчас после овуляции. Прозрачная оболочка содержит примерно 1012 молекул ZP3, вместе с ZP2 формирующих нити длиной 2–3 мкм и толщиной 7 нм (рис. 3-5). В составе этих нитей комплекс ZP2–ZP3 образует структурные повторы каждые 15 нм. Нерегулярным образом нити соединены при помощи ZP1, что и приводит к образованию трёхмерной сети — каркаса прозрачной оболочки. В петлях этого каркаса расположены углеводные компоненты zona pellucida. ZP3 — рецептор сперматозоида. Инактивация или отщепление О-олигосахарида от молекулы ZP3 блокирует связывание сперматозоида с яйцеклеткой. ZP2 — вторичный рецептор сперматозоида.
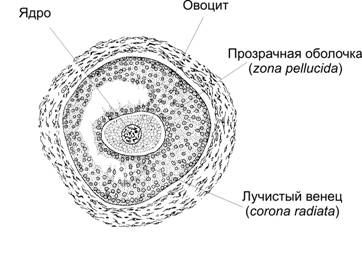
Рис. 3-3. Овоцит. Лучистый венец, прозрачная оболочка и плазматическая мембрана овоцита — барьеры, которые должен последовательно преодолеть сперматозоид, чтобы произошло оплодотворение. [17]
События, предшествующие оплодотворению
Для того, чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера (рис. 3-4) лучистый венец, состоящий из нескольких слоёв фолликулярных клеток, прозрачную оболочку и, наконец, плазматическую мембрану яйцеклетки, при слиянии которой с плазмолеммой сперматозоида и начинается собственно оплодотворение. Сперматозоид легко проникает через лучистый венец между рыхло расположенными фолликулярными клетками и достигает прозрачной оболочки. Прозрачная оболочка — существенный барьер на пути сперматозоида. При взаимодействии сперматозоида с прозрачной оболочкой последовательно происходят следующие события: связывание сперматозоида с его рецептором ® акросомная реакция ® расщепление компонентов zona pellucida ферментами акросомы ® проникновение сперматозоида через образовавшийся в оболочке канал к плазматической мембране яйцеклетки. Взаимодействие сперматозоида с прозрачной оболочкой происходит в два этапа. Сначала сперматозоид слабо прикрепляется к прозрачной оболочке. На этом этапе взаимодействие не видоспецифично, сперматозоиды могут быть легко отделены. На втором этапе сперматозоиды прочно связываются со своими рецепторами. Это взаимодействие видоспецифично (видовую специфичность обеспечивает также акрозин, освобождающийся в ходе акросомной реакции).
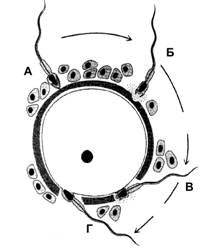
Рис. 3-4. Преодоление сперматозоидом барьеров яйцевой клетки. А — проникновение сперматозоида через лучистый венец; Б — акросомная реакция; В — прохождение сперматозоида через прозрачную оболочку; Г — слияние клеточных мембран сперматозоида и яйцеклетки. [17]
Акросомная реакция
Акросомная реакция — экзоцитоз содержимого акросомы для локального разрушения прозрачной оболочки и преодоления сперматозоидом этого барьера. Начало акросомной реакции ¾ следствие взаимодействия олигосахаридов гликопротеина ZP3 прозрачной оболочки с лектин-подобным рецептором b1,4-галактозилтрансферазой I (GalTI) в мембране головки сперматозоида. При акросомной реакции наружная мембрана акросомы и клеточная мембрана сливаются и формируют мелкие пузырьки (рис. 3-5), отделяющиеся от головки сперматозоида. При этом из акросомы освобождаются гиалуронидаза, протеазы (в том числе акрозин), гликозидазы, липазы, нейраминидаза и фосфатазы. Ферменты расщепляют молекулы прозрачной оболочки, что позволяет сперматозоиду преодолеть этот барьер.
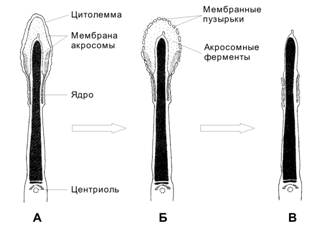
Рис. 3-5. Акросомная реакция. Последовательные этапы акросомной реакции. [17]
Активация G-каскада
Взаимодействие нескольких олигосахаридных остатков ZP3 прозрачной оболочки с GalT I стимулирует агрегацию рецепторов и активацию Gi–белка, что приводит к массированному транспорту внутрь головки сперматозоида Ca2+ и Na+ в обмен на H+. Увеличение внутриклеточной концентрации Ca2+ активирует Ca2+-зависимую фосфолипазу, в результате изменяется уровень вторых посредников — циклических нуклеотидов. Вслед за этим активируется протонная АТФаза, что приводит к увеличению внутриклеточного рН. Увеличение концентрации Ca2+ в цитозоле и повышение pH в головке сперматозоида запускают акросомную реакцию. Как только начинается акросомная реакция, ZP2 дополнительно связывает сперматозоид.
Экзоцитоз
Секреция содержимого акросомы подчиняется правилам Са2+-регулируемого экзоцитоза. В ответ на увеличение концентрации Ca2+ в цитозоле белковый комплекс SNARE (t-SNARE и v-SNARE ) обеспечивает слияние наружной мембраны акросомы и плазмолеммы. Слияние и диссоциацию белков SNARE контролируют ГТФ-связывающий белок Rab3А и АТФ-гидролизующий фермент NSF.
В результате акросомной реакции в прозрачной оболочке образуется узкий канал, через который проходит сперматозоид; плазматические мембраны сперматозоида и яйцеклетки вступают в соприкосновение и сливаются. Вскоре слившиеся мембраны разрушаются, сперматозоид, лишённый собственной плазматической мембраны, оказывается погружённым в цитоплазму яйцеклетки.
Активация яйцеклетки
Оплодотворение активизирует яйцеклетку, в цитоплазме повышается концентрация Ca2+, что служит сигналом для второго деления мейоза. Оплодотворённый овоцит второго порядка после метафазы второго деления завершает мейоз с образованием гаплоидной зрелой яйцеклетки и второго полярного тельца. Оно располагается рядом с первым между прозрачной оболочкой и плазмолеммой.
Овариальная тератома — патология, возникающая в яичниках будущего организма в случае, если второе полярное тельце не выталкивается из зиготы (более того, возможно вхождение его хромосом в состав синкариона — самооплодотворение).
Объединение пронуклеосов
В течение первых 12 часов после проникновения сперматозоида в яйцеклетку происходит перестройка ядер (пронуклеусов) слившихся гамет. Ядра набухают, появляются ядрышки. Пронуклеусы мигрируют в центр яйцеклетки и сближаются. Их ядерные оболочки исчезают, а материнские и отцовские хромосомы перемешиваются — образуется синкарион. Этот процесс (сингамия) и есть собственно оплодотворение. Таким образом, при сингамии происходит объединение ядерного генома гаплоидных гамет: (22 аутосомы + 1 половая хромосома) ´ 2 = 44 аутосомы + 2 половые хромосомы и появляется диплоидная зигота (новый организм, пока одноклеточный). Сперматозоид приносит также митохондриальную ДНК и сигнальные белки дробления — с Mr 14 и 18 кД. Наконец, в ходе оплодотворения происходит завершение мейоза, и детерминируется генетический пол нового организма.
Генетический пол определяет Y-хромосома. Примерно половина сперматозоидов содержит Y-хромосому, другая половина — хромосому Х. Следовательно, половина зигот имеет генетически мужской пол (XY), вторая половина — генетически женский пол (XX).
Митохондриальный геном
Самореплицирующиеся митохондрии имеют собственную ДНК (митохондриальная ДНК), кодирующую полипептиды, частично обеспечивающие функцию этой органеллы. Митохондрии зиготы имеют двоякое происхождение: часть содержалась в овоците (материнская часть митохондриального генома), другую часть этих органелл приносит сперматозоид (отцовский митохондриальный геном).
Наследуемые с митохондриями (отцовскими или материнскими) заболевания (например, врождённая невропатия зрительного нерва Лебера, MERRF- и MELAS–синдромы, дилатационная митохондриальная кардиомиопатия, некоторые скелетномышечные миопатии).
В ходе собственно оплодотворения и сразу после него в зиготе и окружающих её структурах происходит ряд процессов (уменьшение объёма зиготы, формирование перивителлинового пространства, кортикальная реакция, модификация прозрачной оболочки). Назначение этих процессов — предупреждение полиспермии и создание гомеостатической среды для концептуса.
Зигота
При слиянии плазматических мембран гамет и объединении ядерных геномов происходят существенные изменения внутриклеточного ионного состава, приводящие к уменьшению объёма зиготы, деполяризации её плазмолеммы, развитию кортикальной реакции. Прямое следствие уменьшения объёма — образование перивителлинового пространства. Перивителлиновое пространство содержит оба полярных тельца и имеет характерный ионный состав (гомеостатическая среда для концептуса), создающий дополнительное препятствие для проникновения к зиготе других сперматозоидов. Деполяризация плазматической мембраны также предупреждает проникновение сперматозоидов в зиготу.
Кортикальная реакция
По периферии яйцеклетки расположены кортикальные гранулы. Их диаметр 200–600 нм; они содержат ферменты, в том числе различные гидролазы. Сразу после проникновения сперматозоида в яйцеклетку начинается кортикальная реакция — выделение содержимого кортикальных гранул в перивителлиновое пространство. Сигнал для кортикальной реакции — повышение концентрации Ca2+ в цитозоле. Под действием ферментов кортикальных гранул происходят протеолиз ZP2 (ZP2 ® ZP2f) и модификация рецептора сперматозоида ZP3 (ZP3 ® ZP3f). В модифицированном состоянии ZP3f теряет способность к запуску акросомной реакции. Тем самым блокируется полиспермия. Белки SNARE (синтаксин, SNAP-25), регулирующие белки (например, Rab3A) контролируют Са2+-зависимый экзоцитоз кортикальных гранул.
Оболочка оплодотворения
В результате кортикальной реакции прозрачная оболочка подвергается существенным изменениям — происходит её стабилизация (оболочка оплодотворения). В стабилизированном состоянии оболочка оплодотворения защищает проходящий по маточной трубе концептус. Без оболочки оплодотворения дробление зиготы невозможно.
Экстракорпоральное оплодотворение
В акушерско-гинекологической практике часто сталкиваются с невозможностью иметь ребёнка вследствие патологии маточных труб, олигоспермии и т.д. Проблему решают применением экстракорпорального оплодотворения. Сущность метода состоит в получении жизнеспособных яйцеклеток с последующим их оплодотворением in vitro, введением в матку концептуса и возникновением беременности. Если в естественных условиях при овуляции из яичника выходит одна яйцеклетка, то для успешного экстракорпорального оплодотворения необходимо одновременное получение большого их количества. Для этого женщине проводят медикаментозную стимуляцию овуляции гормональными препаратами. Яйцеклетки получают при хирургической операции с применением эндоскопического оборудования или при помощи трансвагинальной техники. Полученные яйцеклетки культивируют in vitro в питательной среде специального состава, где и производят их оплодотворение путём введения в среду сперматозоидов. Дробление контролируют микроскопически, и 4 нормально развивающихся концептуса одновременно вводят через шейку в полость матки. Операция успешна в 20–25% случаев.
Дробление
Дробление — митотическое деление диплоидных клеток без увеличения их суммарного объёма — стадия развития, наступающая после оплодотворения. При дроблении происходит относительно быстрое увеличение количества клеток (бластомеры), от 2 до примерно 12–16 к третьим суткам после оплодотворения, когда концептус достигает стадии морулы и выходит в полость матки из маточной трубы (рис. 3-6).
Сигнальный белок дробления. В зиготе обнаружены два принесённые сперматозоидом белка с Mr 14 и 18 кД, содержащие одну и ту же Аг-детерминанту. АТ к этому Аг блокируют первые дробления зиготы, не оказывая влияния на другие процессы.
Дробление отличается от обычного клеточного цикла отсутствием фаз G1 и G2 и очень короткой S–фазой, во время которой происходит синтез ДНК. Интервал между делениями дробления составляет примерно 12–24 часа. Роста клеток не происходит и в ходе дроблений клетки уменьшаются в размерах, так что диаметр концептуса не превышает 100 мкм. На стадии бластоцисты дробления прекращаются и клетки возобновляют нормальный клеточный цикл G1, S, G2, M.
В ходе дробления и на последующих стадиях развития происходит устранение дефектных клеток путём апоптоза. На всех стадиях развития наблюдается конститутивная экспрессия белка BAX. В ходе развития частота экспрессии проапоптозных и антиапоптозных белков семейства BCL-2 проявляется сходным образом, но на 2-клеточной стадии преобладает экспрессия проапоптозных белков. В отличие от нормальных, клетки дефектных концептусов на стадии 4-х бластомеров экспрессируют BAK.
Характер дробления определяется количеством и распределением в цитоплазме желточных включений. У человека — плацентарного млекопитающего — в связи с внутриутробным питанием плода через плаценту нет необходимости в создании больших запасов желтка. Поэтому желток распределён в цитоплазме равномерно (изолецитальная яйцеклетка). Зигота, образованная такой яйцеклеткой, совершает голобластическое дробление, т.е. полностью разделяется на два бластомера. Дальнейшие дробления — асинхронные и несколько неравномерные. При дроблении зигота окружена прозрачной оболочкой, сохраняющейся до стадии бластоцисты.

Рис. 3-6. Концептус от оплодотворения до имплантации. Овуляция наступает на 14-й день менструального цикла, в течение 1 сут происходит оплодотворение. Трое суток с момента оплодотворения концептус продвигается по маточной трубе, на 4 сут попадает в полость матки и через 5,5–6 сут имплантируется в эндометрий. В ходе перемещения по маточной трубе происходит дробление, и в полости матки оказывается бластоциста, готовая к имплантации. 1 — овоцит тотчас после овуляции, 2 — оплодотворение через 12–24 ч после овуляции, 3 — стадия появления мужского и женского пронуклеусов, 4 — начало дробления, 5 — двухклеточная стадия (30 часов), 6 — морула, состоящая из 12–16 бластомеров (3 сут), 7 — продвинутая морула в полости матки (4 сут), 8 — ранняя бластоциста (4,5–5 сут), 9 — имплантация бластоцисты (5,5–6 сут). [17]
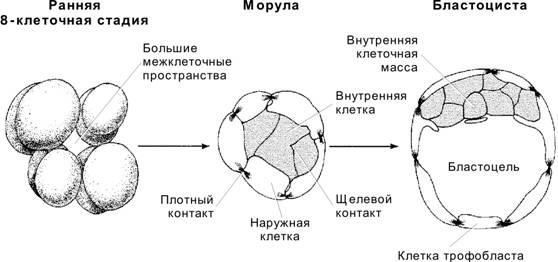
Рис. 3-7. Компактизация и морула на ранних этапах развития. В результате компактизации уменьшается размер межклеточных пространств между бластомерами, они сближаются, и формируется морула. В моруле различают внутреннюю часть (клетки связаны щелевыми контактами) и наружную часть (клетки соединены при помощи плотных контактов). Из клеток внутренней части морулы развивается эмбриобласт, а из клеток наружной части формируется трофобласт. Внутренняя клеточная масса и трофобласт — главные структурные элементы бластоцисты. [17]
Первое дробление (рис. 3-6) начинается примерно через 30 часов после оплодотворения. Плоскость первого деления проходит через область направительных телец. Поскольку желток в зиготе распределён равномерно, выделение анимального и вегетативного полюсов затруднено. Область отделения направительных телец обычно рассматривают как анимальный полюс. Образующиеся два бластомера несколько различны по размерам.
В результате первого деления дробления нарушается симметрия зародыша и образуется два бластомера с различными потенциями развития: один бластомер дает начало «эмбриональной части» бластоцисты (полярный трофобласт и глубокая часть внутренней клеточной массы), а другой бластомер — «неэмбриональной части» бластоцисты (муральный трофобласт и поверхностный слои внутренней клеточной массы).
Второе дробление (рис. 3-6). Образование второго митотического веретена в каждом из бластомеров происходит вскоре после окончания первого деления, плоскость второго деления проходит перпендикулярно плоскости первого деления дробления, концептус переходит в стадию 4-х бластомеров. Однако, дробление у человека асинхронно, поэтому при переходе от стадии 2-х бластомеров к стадии 4-х бластомеров в течение некоторого времени можно наблюдать трёхклеточный концептус. На стадии 4-х бластомеров синтезируются все основные типы РНК.
Третье дробление. На этой стадии несинхронность дробления проявляется в большей мере, в итоге образуется концептус с различным количеством бластомеров; условно его можно отнести к стадии 8 бластомеров (рис. 3-6, рис. 3-7). До этого бластомеры расположены рыхло. Непосредственно перед следующим (4-м) делением дробления с образованием 16 бластомеров эмбрион подвергается компактизации (рис. 3-8). Концептус уплотняется (компактизация), поверхность соприкосновения бластомеров увеличивается, объём межклеточного пространства уменьшается. Сближение бластомеров в ходе компактизации — необходимое условие для образования между бластомерами специализированных межклеточных контактов (щелевых и плотных), формирующихся на поздней 8-клеточной стадии (морула). До формирования контактов в плазматическую мембрану бластомеров начинает встраиваться E-кадгерин (увоморулин) — белок адгезии клеток. В бластомерах ранних концептусов E-кадгерин равномерно распределён в клеточной мембране. Позднее в области межклеточных контактов образуются скопления (кластеры) молекул E-кадгерина.
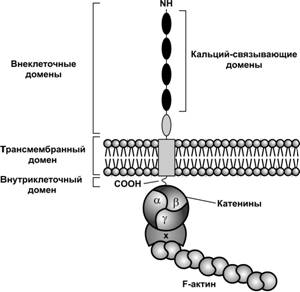
Рис. 3-8. Молекула E-кадгерина в бластомере связана с примембранным цитоскелетом (актином) через катенин. Клетки начинают экспрессировать Ca2+-зависимый белок адгезии E-кадгерин, который способствует сближению (компактизации) и поляризации клеток. [102]
В ходе компактизации E-кадгерин способствует поляризации клеток. Между апикально-латеральными областями наружных бластомеров формируются плотные контакты, достигающие максимального развития на 32-клеточной стадии, когда начинает формироваться полость (бластоцель). Позднее, в ходе гаструляции, E-кадгерин замещается N–кадгерином.
Морула
По мере увеличения числа бластомеров концептус перемещается по маточной трубе и к третьим суткам достигает стадии морулы (рис. 3-6). Морула — группа клеток, возникших в ходе нескольких делений дробления и заключённых внутри прозрачной оболочки (рис. 3-7). Центрально расположенные клетки морулы образуют щелевые контакты, при помощи которых осуществляются информационные межклеточные взаимодействия. Именно из этих клеток и возникает собственно эмбрион. Периферические клетки морулы соединяют плотные контакты; они формируют барьер, обособляющий внутреннюю среду морулы.
Для внутренних клеток морулы характерна выраженная экспрессия Nanog. LIF/STAT3-независимый фактор транскрипции Nanog необходим для самоподдержания эмбриональных стволовых клеток и сохранения их плюрипотентности. В ходе дальнейшего развития nanog (ген плюрипотентности) экспрессируется клетками внутренней клеточной массы бластоцисты. По мере развития экспрессия nanog снижается и сохраняется только в первичных половых клетках.
В обособлении в моруле клеток будущей внутренней клеточной массы также участвует фактор транскрипции Oct4, при этом происходит подавление экспрессии генов, ответственных за дифференцировку клеток в трофобласт. Oct4, как и Nanog, отвечает за поддержание плюрипотентности эмбриональных стволовых клеток и в дальнейшем Oct4 также экспрессируется клетками внутренней клеточной массы.
С другой стороны, на стадии поздней морулы поляризованные бластомеры наружного слоя экспрессируют cdx2 (caudal-related homeobox gene). Повышенная экспрессия фактора транскрипции Cdx 2 индуцирует формирование трофобласта. Гены Cdx1 и Cdx2, кодирующие соответствующие факторы транскрипции, гомологичны гену, содержащему гомеобокс, Caudal у Drosophila. Cdx 1 экспрессируется на ранних стадиях развития, когда определяются оси тела; на более поздних стадиях экспрессия Cdx 1 характерна только для клеток эмбриональной энтодермы, дающих начало кишечной трубке. У взрослого человека Cdx 1 экспрессируется стволовыми эпителиальными клетками крипт кишечника.
Бластоциста
Бластоциста возникает с появлением бластоцеля (заполненной жидкостью полости) к четвёртым суткам после оплодотворения (рис. 3-7, рис. 3-9, рис. 3-21А). Объём бластоцеля увеличивается, и концептус приобретает форму пузырька. Прозрачная оболочка истончается и исчезает. Если не произойдёт растворения прозрачной оболочки, бластоциста не сможет прикрепиться к поверхности эндометрия. Бластоцисту составляют трофобласт и внутренняя клеточная масса (эмбриобласт).
Трофобласт — покров зародышевого комплекса (рис. 3-7, рис. 3-9); образован периферическими клетками морулы, связанными плотными контактами. Крупные удлинённые клетки трофобласта соединены при помощи плотных контактов, что стабилизирует состав жидкости в бластоцеле. Клетки трофобласта «накачивают» жидкость в бластоцель.
Внутренняя клеточная масса (эмбриобласт) — компактная масса мелких клеток, выступающих в бластоцель (рис. 3-7, рис. 3-9). Если трофобласт образуется относительно быстро делящимися клетками, то внутренняя клеточная масса представлена клетками с меньшей частотой делений. Клетки внутренней клеточной массы происходят из центральной части морулы и связаны при помощи щелевых контактов. В дальнейшем из внутренней клеточной массы образуются собственно зародыш и некоторые связанные с ним оболочки. Частичное или полное разделение внутренней клеточной массы приводит к развитию близнецов.
Гаструляция
Первичные зародышевые листки
Гаструляция начинается в конце второй недели развития и характеризуется появлением у клеток способности к перемещениям. Необходимо иметь в виду, что не существует резкой временной границы между бластоцистой (рис. 3-9) и гаструлой. С началом гаструляции активируются первые тканеспецифические гены. Эмбриобласт расслаивается на эпибласт (слой цилиндрических клеток) и гипобласт (слой кубических клеток, обращённый к бластоцелю). Эпибласт и гипобласт вместе образуют двухслойный зародышевый диск (бластодиск). В дальнейшем на месте двухслойного зародышевого диска путём миграции и пролиферации клеток развиваются первичные зародышевые листки: эктодерма, мезодерма и энтодерма. Гаструляцию можно определить как морфогенетический процесс, в ходе которого из однослойного эпибласта образуются три зародышевых листка: энтодерма, мезодерма и эктодерма.
Гипобласт
Формирование гипобласта (первичной энтодермы) происходит по каудально-краниальному градиенту. Обращённые к бластоцелю клетки вентральной части внутренней клеточной массы обособляются в тонкий слой — гипобласт (рис. 3-9). Клетки гипобласта выселяются из внутренней клеточной массы вследствие слабого адгезионного взаимодействия между ними. Интенсивно пролиферирующие клетки гипобласта перемещаются по внутренней поверхности трофобласта и формируют внезародышевую энтодерму прилегающей к трофобласту стенки желточного мешка.
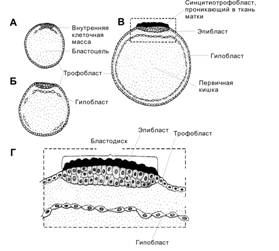
Рис. 3-9. Начало гаструляции. Стадии формирования первичной кишки (А–В), бластодиск — В и Г. Выселяющиеся из гипобласта клетки обрастают изнутри трофобласт, формируя внезародышевую энтодерму. [17]
Эпибласт
Оставшиеся после образования гипобласта клетки внутренней клеточной массы формируют эпибласт (первичная эктодерма). Из него развивается зародыш, а также внезародышевые мезодерма и эктодерма (амниотическая).
Зародышевый диск
Зародышевый диск (рис. 3-9, рис. 3-10) — основная структура, участвующая в гаструляции. Зародышевый диск (более ранняя стадия — бластодиск) состоит из эпибласта, находящегося в контакте с гипобластом (фактически уже слоем клеток внезародышевой энтодермы желточного мешка). Краниальный отдел зародышевого диска расширен. В дальнейшем в суженной каудальной части зародышевого диска возникает вытянутое в кранио-каудальном направлении утолщение — первичная полоска. Первичная полоска появляется на 14-е сутки и чётко различима с 15–16-х суток развития. Через первичную полоску мигрируют клетки, участвующие в образовании первичных зародышевых листков и внезародышевых структур (рис. 3-10, рис. 3-12).
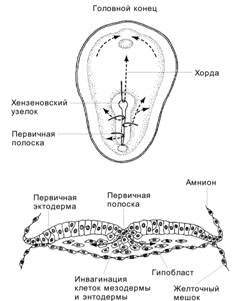
Рис. 3-10. Перемещения клеток при гаструляции. Клетки первичной эктодермы, проходящие через гензеновский узелок, образуют хорду. Остальные клетки первичной эктодермы, проходящие через первичную полоску, мигрируют в латеральном направлении и формируют мезодерму и энтодерму. [17]
Зародышевая энтодерма происходит из части эпибласта, расположенной кпереди от первичной полоски.
Зародышевая и внезародышевая мезодерма. Через заднюю часть первичной полоски мигрируют клетки эпибласта, образующие мезодерму. Сначала перемещаются клетки будущей внезародышевой мезодермы, а затем клетки для мезодермы зародыша. Хорду формируют мезодермальные клетки, расположенные вдоль срединной линии зародыша. Они мигрируют через головной конец первичной полоски в области гензеновского узелка по направлению к будущей головной части зародыша.
Внезародышевая эктодерма. Из периферических участков эпибласта выселяются клетки, выстилающие изнутри полярный трофобласт.
Эктодерма зародыша образуется за счёт не выселяющихся из эпибласта клеток.
Молекулярные механизмы гаструляции
Третья неделя развития характеризуется установлением полярности (осей): кранио-каудальной, дорсо-вентральной и «правое-левое». Гены, контролирующие закладку головы, сначала экспрессируются в краниальной части гипобласта — передней висцеральной эктодерме. Они кодируют синтез транскрипционных факторов OTX2, LIM1 и HESX1. После спецификации краниального отдела гипобласта начинается формирование первичной полоски в каудальном его отделе. Полоска индуцируется фактором Nodal, принадлежащем надсемейству TGFb. При этом одновременно с установлением кранио-каудальной оси формируется дорсо-вентральная. Другой член надсемейства трансформирующего фактора роста b (TGFb), морфогенетический белок кости 4 (BMP4), секретируемый клетками зародышевого диска, вместе с фактором роста фибробластов (FGF) поддерживает дифференцировку мезодермы и образование промежуточной и латеральной её частей. В клетках гензеновского узелка активируется выработка ноггина, фоллистатина и фактора goosecoid, которые ингибируют BMP4. Это инициирует спецификацию мезодермы по дорсо-вентральной оси (дорсализацию) и формирование нотохорда, параксиальной мезодермы (сомитомеров) и сомитов. Исключительно важным фактором для процесса гаструляции считается FGF8. При отсутствии FGF8 клетки эпибласта мигрируют в область полоски, но утрачивают способность впоследствии выселяться из неё, что исключает возможность формирования энтодермы и мезодермы. При этом отсутствие мезодермы делает невозможной спецификацию дорсальной эктодермы и формирование осевых структур. Вырабатываемый в узелке FGF8 инициирует экспрессию транскрипционного фактора Nodal в мезодерме левой половины зародыша. В свою очередь Nodal активирует сигнальный каскад с участием lefty2 и транскрипционного фактора PITX2 (рис. 3-11). Поскольку узелок служит источником многочисленных факторов (ноггин, фоллистатин, goosecoid, FGF8 и др.), определяющих процессы закладки зародышевых листков и их последующую спецификацию, его считают ключевой структурой и называют «организатором». Термин «организатор» употребляют применительно к зачаткам органов живых зародышей, продуцирующих индукторы.
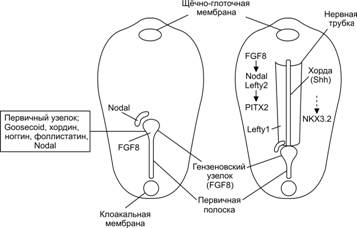
Рис. 3-11. Молекулярные события на стадии гаструляции. Гензеновский узелок как организующая структура служит источником ноггина, хордина, фоллистатина, факторов goosecoid и Nodal. Два последних вместе с фоллистатином угнетают эффекты морфогенетического белка кости 4 (BMP4), который равномерно секретируется по всему зародышевому диску и вместе с FGF поддерживает спецификацию мезодермы на промежуточную и латеральную. Секреция FGF8 в области узелка запускает каскад событий, приводящих к установлению полярности «правое–левое». Этот фактор индуцирует экспрессию транскрипционного фактора Nodal в мезодерме на левой стороне с последующим усилением экспрессии последовательно факторов Lefty2 и PITX2. На правой стороне этот каскад не запускается, т.к. здесь он сдерживается совместным действием экспрессируемого на левой стороне нервной пластинки фактора lefty1 и Shh из хорды. Транскрипционный фактор NKX3.2 — наиболее вероятный кандидат на роль молекулярного сигнала, экспрессируемого на правой стороне и поддерживающего её спецификацию. [112]
Правое–левое
Экспрессия lefty1 на левой стороне нервной пластинки, которая возникнет на последующей стадии нейруляции, вместе с действующим из нотохорда Sonic hedgehog (Shh) оказывает совместное влияние на дифференцировку структур на правой стороне, блокируя экспрессию молекул, характерных для левой стороны. Гены, контролирующие специфику развития правой стороны, пока не установлены. Возможным кандидатом служит ген, кодирующий транскрипционный фактор NKX3.2 (рис. 3-11). Нарушение молекулярного механизма, кодирующего специфику структур на правой и левой стороне, приводит к situs inversus (сердце справа, печень слева и т.д.).
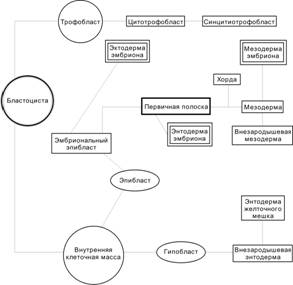
Рис. 3-12. Происхождение зародышевых листков. [17]
Этические проблемы. Две недели пренатального развития человека, срок формирования первичной полоски, рассматривают как своеобразную критическую точку для последующей нейруляции и формирования нервной системы. Именно поэтому многие эмбриологи, руководствуясь соображениями этического порядка, предлагают этим периодом ограничить использование для научных исследований эмбрионов человека, полученных методом экстракорпорального оплодотворения. Хотя существующие в этом плане научные программы имеют целью выяснение важных вопросов генеза наследственных болезней человека, причин бесплодия и т.д. Репродуктивная технология столкнулась со множеством моральных и юридических проблем, решением которых занимаются специальные группы экспертов по этике экстракорпорального оплодотворения и трансплантации эмбрионов человека.
На сегодняшний день трудности получения экспериментального материала и этические нормы ограничивают наши знания в области раннего эмбриогенеза у человека. По этой причине основные положения раннего эмбриогенеза приведены на примере развития мыши — наиболее изученного млекопитающего.
Нейруляция, сомиты, органогенез
В результате гаструляции сформировались зародышевые листки, части которых, влияя друг на друга, индуцируют образование новых структур. Пример подобного влияния — первичная эмбриональная индукция; её результат — развитие из дорсальной эктодермы нервной системы (ткани). Нейруляция — процесс закладки нервной системы и осевых структур. Нейруляция начинается с 16 суток развития (первые признаки формирования нервной пластинки) и в основном завершается к 22–23 суткам. Почти одновременно из мезодермы формируются сомиты и нефротом.
Стадии нейруляции (рис. 3-13) — индукция (первичная эмбриональная индукция) нервной пластинки ® приподнимание краёв нервной пластинки и образование нервного желобка ® появление нервных валиков ® формирование нервного гребня и начало выселения из него клеток ® смыкание нервных валиков с образованием нервной трубки ® срастание эктодермы над нервной трубкой. Некоторые структуры нервной ткани развиваются из нейрогенных плакод.
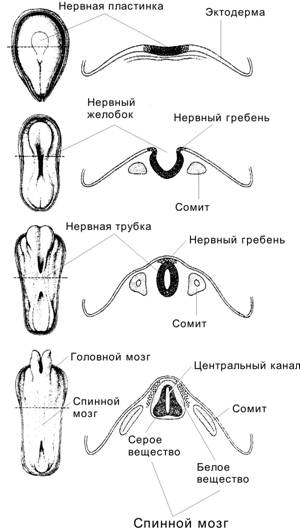
Рис. 3-13. Развитие нервной системы. Последовательные стадии (сверху вниз): нервная пластинка (19 сут), нервный желобок (20 сут), нервная трубка (22 сут) и зачаток ЦНС (23–24 сут). Слева — дорсальная поверхность зародыша, справа — дорсальная часть зародыша в поперечном разрезе на уровне, обозначенном пунктиром слева. [17]
Первичная эмбриональная индукция
Нейральная, или первичная эмбриональная индукция — образование нервной пластинки из дорсальной эктодермы. Этот процесс определяет организатор — хордомезодерма. В ходе первичной эмбриональной индукции детерминируется судьба клеток, дающих начало нервной системе. Природа индуктора и механизм индукционного взаимодействия между хордомезодермой и дорсальной эктодермой неясны. Возможно, клетки хордомезодермы выделяют химический фактор, вызывающий индукцию. Это гипотетическое вещество называют нейрализующим фактором. В этом случае отношения между хордомезодермой и дорсальной эктодермой уместно рассматривать в рамках концепции “сигнал ® мишень”. С другой стороны, получены данные о роли программы самих эктодермальных клеток в образовании зачатка нервной системы. Так, на ранних эмбрионах шпорцевой лягушки показано, что ещё до вступления в контакт с мезодермой эктодермальные клетки уже предрасположены к дифференцировке в нервную ткань. Начало дифференцировки клеток развивающейся нервной пластинки контролируют гены, содержащие гомеобокс. Вероятно, что ещё до гаструляции клетки дорсальной эктодермы предетерминированы для превращения в нервную ткань.
Нервная пластинка
Нервная пластинка — утолщённая часть дорсальной эктодермы, формирующаяся по кранио-каудальному градиенту. При нейруляции большое значение имеет изменение формы клеток дорсальной эктодермы: при формировании нервной пластинки происходит удлинение клеток, при этом микротрубочки в них ориентируются параллельно дорсо-вентральной оси. Призматические клетки только что сформированной нервной пластинки расположены на базальной мембране, содержащей фибронектин, сульфатированные гликозаминогликаны и ламинин. Клетки нервной пластинки в апикальной части соединены при помощи плотных контактов, а в базальной части — щелевых.
Щелевые контакты и образование нервной пластинки. В формировании нервной пластинки, а также в её дальнейшей дифференцировке важную роль играют информационные взаимодействия между клетками через щелевые контакты. Так, обработка зародышей АТ против белков щелевых контактов предотвращает формирование нервной пластинки.
Нервная трубка
Вскоре после образования края нервной пластинки приподнимаются, и формируются нервные валики (рис. 3-13). Между валиками расположен нервный желобок. Позднее края нервных валиков смыкаются по срединной линии, и образуется замкнутая нервная трубка. Краниальный и каудальный участки нервной трубки долго остаются незамкнутыми, их называют соответственно передним и задним нейропором. Передний нейропор закрывается на 23–26-й день развития, а задний — на 26–30-й день.
Нервный гребень
После смыкания валиков и образования нервной трубки часть эктодермы, расположенная между нейральной и ненейральной (кожной) эктодермой (рис. 3-13), формирует новую структуру — нервный гребень, его производные см. в таблице 3-3.
Таблица 3-3. Производные нервного гребня
|
Чувствительные нейроны спинномозговых узлов и ганглиев черепных нервов: верхнего ганглия языкоглоточного нерва, яремного ганглия, часть нейронов ганглия тройничного нерва, часть нейронов ганглия коленца промежуточного нерва |
|
Симпатические нейроны |
|
Парасимпатические нейроны |
|
Шванновские клетки и клетки-сателлиты спинномозговых узлов и ганглиев черепных нервов |
|
Меланоциты |
|
Клетки каротидного тельца |
|
Продуцирующие кальцитонин клетки |
|
Хромаффинные клетки |
|
Хрящ, кости, мышцы и соединительная ткань лица |
|
Окологлоточная мезенхима |
|
Верхне- и нижнечелюстной отростки |
|
Подъязычные дуги и 3-я глоточная дуга |
|
Одонтобласты |
|
Эндотелий роговицы |
|
Клетки в стенке дуги аорты |
Нейрогенные плакоды
Нейрогенные плакоды — утолщения эктодермы, расположенные латерально по обе стороны от формирующейся нервной трубки в краниальном отделе зародыша. Производные нейрогенных плакод — нейроны обонятельной выстилки, нейроны вестибулярного и слухового ганглиев, а также чувствительные нейроны коленчатого, каменистого, узловатого и тройничного ганглиев черепных нервов.
Молекулярные сигналы нейруляции
На стадии нервной пластинки на 3-й неделе развития все клетки пластинки экспрессируют гомеобокс–содержащие гены Pax3 и Pax7 (рис. 3-14). Нотохорд служит источником фактора Shh, который ингибирует экспрессию генов Pax3 и Pax7 в клетках средней и вентральной части нервного желобка, начиная с момента его образования. Мутации Shh приводят к утрате всех вентральных срединных структур, включая дно желобка, а также мотонейронов. Кроме Shh, в нотохорде вырабатываются антагонисты BMP — ноггин, хордин и фоллистатин. Из них только ноггин экспрессируется в области дна пластинки, что свидетельствует о его непосредственной роли в формировании нервной трубки. К выработке фактора Shh подключаются клетки вентральной части нервной трубки. Ингибирование экспрессии генов pax3 и pax7 вместе с нарастанием концентрации Shh детерминирует процесс «вентрализации» нервной трубки с последующей закладкой в вентральной её части мотонейронов. В дорсальной части нервного желобка, а позднее и нервной трубки экспрессия генов pax3 и pax7 не угнетается как в вентральной части, а наоборот поддерживается, в чём участвуют BMP4 и BMP7, источником которых служит соседняя «ненервная» эктодерма (рис. 3-15).
Дорсалин-1 принадлежит надсемейству трансформирующего фактора роста b (TGFb). Избирательно экспрессируется в дорсальной части нервной трубки, начиная с момента её смыкания, и поддерживает здесь дифференцировку клеток. На последующих стадиях развития спинного мозга присутствует преимущественно в дорсо-медиальной части зачатка. Сегментарную спецификацию нервной трубки контролируют гены Hox/Lim.

Рис. 3-14. Молекулярные сигналы нейруляции. На третьей неделе развития во всей нервной пластинке отмечена экспрессия гомеобокс-содержащих генов PAX3 и PAX7. На этапе формирования нервных желобков секретируемый сначала хордой, а затем вентральной частью нервной трубки Shh сдерживает экспрессию PAX3 и PAX7 в вентральной части нервной трубки. Экспрессию PAX3 и PAX7 в дорсальной части нервной трубки поддерживают морфогенетические белки кости (BMP4 и BMP7) из ненейрогенной эктодермы. [112]
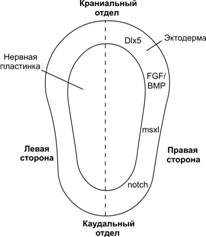
Рис. 3-15. Детерминация границы между нервной пластинкой и окружающей эктодермой. Ростральную границу определяет ген Dlx5, боковую границу детерминируют продукты генов FGF/BMP и MSX1. Строго в пределах нервной пластинки экспрессируется ген Notch. [126]
Дефекты нейруляции
Известно большое число разнообразных дефектов нейруляции, например, грыжи (энцефалоцеле, менингомиелоцеле), нейрокристопатии, аномалии сердца. Дефекты формирования нервной трубки, а также дефекты выселения клеток из нервного гребня и их последующей миграции (нейрокристопатии) встречаются у 1–2 на 1000 новорождённых. В этих случаях в амниотической жидкости плода возрастает содержание a-фетопротеина и ацетилхолинэстеразы.
Пороки развития нервной трубки
Craniorachischisis totalis — редко встречающийся дефект — незаращение нервной трубки.
Анэнцефалия — отсутствие большого мозга.
Миеломенингоцеле — выпадение мозговых оболочек и мозгового вещества вследствие неполного закрытия дужек позвонков. При менингомиелоцеле образуется грыжевое выпячивание через дефекты позвоночного столба (например, spina bifida). Этот порок чаще возникает в пояснично-крестцовом отделе позвоночника. В результате нарушения нормального формирования позвоночного столба происходит выбухание спинного мозга с потерей всех функций ниже уровня поражения. Частое осложнение такого порока — гидроцефалия.
Энцефалоцеле — грыжа головного мозга.
Нейрокристопатии — результат нарушения миграции, дифференцировки и межклеточных взаимодействий производных нервного гребня. Они часто сочетаются и проявляются в виде расщепления века, радужной оболочки или дефекта хрусталика, аномалий сердца, задержки физического и умственного развития, гипоплазии гениталий в мужском организме, аномалий органа слуха и глухоты.
Врождённый мегаколон. Пример нарушения развития производных нервного гребня — болезнь Хиршспрунга.
Аномалии сердца. Клетки нервного гребня мигрируют в сердце и входят в состав некоторых его частей. При искажениях миграции появляются дефекты верхней части межжелудочковой перегородки, а также возникают смещение выносящего сосуда правого желудочка в область межжелудочковой перегородки и незаращение артериального (боталлова) протока.
Клетки зародышевой мезодермы выселяются из эпибласта и формируют пресомитную мезодерму, из которой возникают сомиты — симметричные парные структуры по бокам от хорды и нервной трубки (рис. 3-16). Из мезодермы образуется ещё два крупных зачатка: нефротом (промежуточная мезодерма) и латеральная мезодерма. Производные этих структур, а также всех зародышевых листков см. на рисунке 3-12.
Пресомитная мезодерма
Клетки, прошедшие через первичную полоску, мигрируют в латеральном направлении и образуют непрерывный пласт толщиной в несколько клеток. В непосредственной близости от нервной трубки и хорды мезодермальные клетки образуют скопление — концентрические слои клеток метамерной организации в виде потенциальных сомитов, или сомитомеров, появляющихся очень рано — во время гаструляции. Сомитомеры парааксиальной мезодермы определяют сегментацию хорды, нервной трубки, промежуточной и латеральной мезодермы (мезодермы латеральной пластинки).
Сомиты
В результате пролиферации клеток, их миграции и последующей агрегации из сомитомеров формируется дорсальная мезодерма — сомиты (рис. 3-16). Образование сомитов происходит от головного к хвостовому концу зародыша параллельно с регрессией гензеновского узелка. Новая пара сомитов образуется кзади от последней уже сформированной пары через определённый промежуток времени. Этот интервал составляет в среднем 6,6 часа. В сомите существует полость, ограниченная клетками, связанными между собой при помощи плотных контактов. В каждом сомите различают склеротом, дерматом и миотом; их клетки имеют свои пути миграции и служат источником для различных структур.
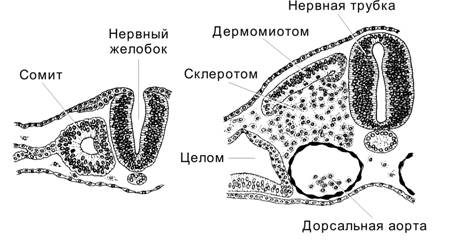
Рис. 3-16. Образование сомита и последующее выселение из него клеток. Слева — мезодермальные клетки сосредоточены латеральнее нервного желобка вокруг небольшой полости; справа — клетки вентральной и медиальной части сомита, расположенного латеральнее нервной трубки, начинают мигрировать в направлении хорды; совокупность этих клеток — склеротом. [17]
Склеротом. Под влиянием хорды и нервной трубки клетки вентро-медиальной области сомитов (склеротом) интенсивно размножаются и выселяются из сомита, окружая хорду и вентральную часть нервной трубки. Выселившиеся клетки дифференцируются в хрящевые и образуют позвонки, рёбра и лопатки.
Миотом и дерматом. В оставшейся дорсо-латеральной части сомита выделяют миотом (внутренний слой клеток, образующий впоследствии скелетную мускулатуру) и дерматом (наружный слой, зачаток соединительнотканной части кожи).
Молекулярные сигналы дифференцировки сомита. Секреторный сигнал Shh действует из вентральной части нервной трубки и нотохорда и индуцирует экспрессию транскрипционного фактора Pax1 в клетках зачатка склеротома (рис. 3-17). В дальнейшем эти клетки приобретают способность дифференцироваться в остеогенном направлении. Активация BMP4 в дорсальной эктодерме вызывает выделение факторов Wnt из дорсальной части нервной трубки, а также из эктодермы, непосредственно прилежащей к сомиту. В первом случае Wnt индуцирует экспрессию мышечно-специфического гена myf5 в дорсо-медиальной части сомита, а во втором — другого мышечно-специфического гена MyoD в вентро-латеральной его части. Под влиянием этих генов клетки дорсо-медиальной части сомита образуют скелетную мускулатуру спины, а вентромедиальной части — остальные мышцы тела и конечностей. Развитие соединительнотканной части кожи контролирует транскрипционный фактор Pax3, который определяет спецификацию клеток сомита, расположенных в средней его части. Сигналом для индукции Pax3 служит нейротрофин 3 (NT3), действующий из дорсальной части нервной трубки.
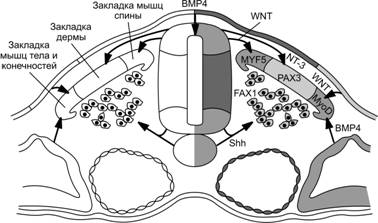
Рис. 3-17. Источники и мишени молекулярных сигналов дифференцировки сомита. [112]
Нефротом
Латеральнее сомита промежуточный листок представлен скоплением клеток промежуточной мезодермы (нефротом) — зачаток мочевыделительной и половой систем.
Латеральная мезодерма
Расположенная латеральнее нефротома мезодерма (мезодерма латеральной пластинки) расщеплена на два листка: дорсальный и вентральный. Дорсальный (париетальный) листок — соматическая мезодерма (из неё образуются серозные оболочки). Вентральный (висцеральный) листок — спланхническая мезодерма (из неё образуются сердце, кора надпочечников, строма гонад, соединительная и гладкомышечная ткани внутренних органов и кровеносных сосудов).
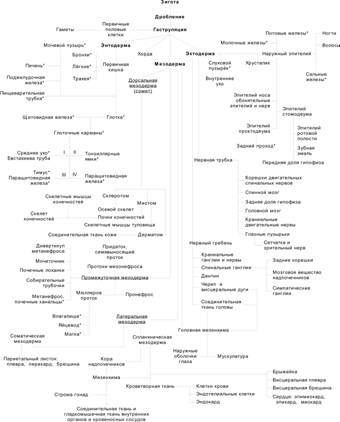
Рис. 3-18. Производные зародышевых листков. [17]
На 4-й неделе завершается нейруляция, начинается активная закладка органов — органогенез. На этом сроке появляются зачатки конечностей и закладываются основные системы органов (рис. 3-18, 3-19), но процесс их роста и становления функций продолжается в плодном и постнатальном периодах. Согласно клональной теории развития, любая ткань и орган берут начало из небольшой группы клонов, каждый из которых образуется из своей стволовой клетки. Например, фоторецепторные клетки обоих глаз формируются из 20 клонов, проксимальные канальцы почек происходят из 4–5 клеток. На ранних стадиях становления общего плана тела важную роль играет мезодерма, служащая носителем позиционной информации. Решающее значение в органогенезе имеют индукционные взаимодействия между клетками.
Формирование целого органа осуществляется в ходе сложных морфогенетических процессов, которые, в свою очередь, основываются на молекулярно-генетических событиях. Участвующие в формировании целого органа молекулярно-морфогенетические системы запускаются несколькими или даже одним геном. Активация этих генов приводит к включению ткане- и органоспецифических синтезов, определяющих специфику данного органа.
Дефекты органогенеза. Эмбрион в периоде органогенеза наиболее чувствителен к действию тератогенных факторов. Большинство врождённых пороков возникает именно в этот критический период.
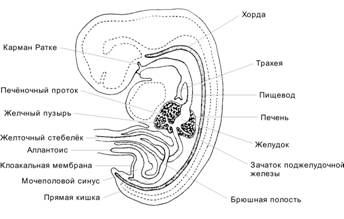
Рис. 3-19. Шестинедельный эмбрион. Сформированы зачатки всех основных систем. В пищеварительной системе на этом сроке расширяется область желудка, закладывается печень, и формируются оба зачатка поджелудочной железы. [17]
Развитие конечностей
Закладка конечностей из материала боковой поверхности тела начинается в середине 4-й недели и заканчивается после формирования пальцев к 8-й неделе. Верхние конечности на 1,5–2 суток опережают закладку нижних. На ранней стадии зачаток конечности представлен преимущественно мезенхимой (рис. 3-20), которая происходит из соматической части латеральной мезодермы. Снаружи мезодермальный зачаток покрыт эктодермой. Морфогенез конечности, как и любого другого органа, целесообразно рассматривать в пространственно-временных координатах. В ходе развития морфогенетическое поле зачатка преобразуется одновременно по трём осям: проксимо-дистальной, дорсо-вентральной и передне-задней. Морфогенез зачатка на каждом конкретном этапе, а также по каждой конкретной оси контролируют различные морфогены. Очень быстро эктодермальные клетки в области кончика зачатка начинают размножаться, вследствие чего эктодермальный покров в этом месте утолщается, формируя апикальный эктодермальный гребень. Его клетки оказывают индуцирующее влияние на клетки прилежащей мезенхимы, инициируя их пролиферацию. Группа активно пролиферирующих мезенхимных клеток, прилежащая к апикальному эктодермальному гребню, названа зоной прогресса. Именно она поддерживает процесс удлинения конечности в проксимо-дистальном направлении. Интенсивность пролиферации клеток в проксимальной части зоны прогресса снижается, затем они останавливают свой рост и формируют компактные скопления, которые дифференцируются в хрящевую ткань. В дальнейшем на основе этого хряща будут сформированы кости конечности по проксимо-дистальному градиенту, т.е. сначала плечо, потом предплечье, далее кости кисти и, наконец, кости пальцев. Таким образом, апикальный эктодермальный гребень регулирует рост конечности по проксимо-дистальной оси. Кроме апикального эктодермального гребня в морфогенетическом поле конечности формируется ещё один регулятор развития — зона поляризующей активности. Она контролирует спецификацию частей конечности по передне-задней оси, что можно пояснить на примере развития пальцев. Применительно к ним зона поляризующей активности необходима для того, чтобы обеспечить развитие большого пальца в лучевой (передней) части зачатка, а закладку мизинца «разрешить» в локтевой (задней) его части, а не наоборот. В эксперименте трансплантация дополнительной зоны поляризующей активности в зачаток конечности дуплицирует пальцы.
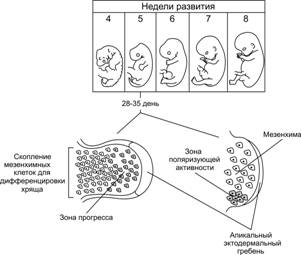
Рис. 3-20. Развитие конечностей у эмбриона на разных сроках (А). Часть эктодермального покрова в области кончика зачатка формирует апикальный эктодермальный гребень (Б). Прилежащая к нему мезенхима образует зону прогресса, которая обеспечивает удлинение конечности по проксимо-дистальной оси. Зона поляризующей активности контролирует развитие конечности по передне-задней оси. [112]
Молекулярные сигналы в развитии конечности
Спецификацию развития конечности определяют гомеозисные гены Hox. Формирование зачатка конечности инициируют FGF8, FGF10, которые продуцируются и секретируются мезенхимными клетками до формирования зачатка. В эксперименте с иммобилизованным на микросферах (мельчайших частицах из искусственного полимера) FGF8 показано образование эктопических конечностей.
Формирование апикального эктодермального гребня индуцируют представители семейства BMP, секретируемые эктодермальными клетками передней части зачатка.
Проксимо-дистальная ось. Формирование апикального эктодермального гребня индуцирует фактор Wnt7a. В ответ на действие этого сигнала клетки апикального эктодермального гребня секретируют FGF2, FGF4 и FGF8, которые стимулируют пролиферацию мезенхимных клеток в зоне прогресса и удлинение зачатка по проксимо-дистальной оси. Дифференцировка клеток по этой оси зависит от времени их пребывания в зоне прогресса, контролируемой 9–13 гомеозисными генами Hox A и Hox D.
Дорсо-вентральная ось. Соматическая мезодерма служит источником транскрипционного фактора En1 (Engrailed-1), который действует на окружающую мезенхиму, а от неё индуцирующий сигнал передаётся на прилежащую эктодерму. Далее эктодермальные клетки дорсальной поверхности зачатка начинают секретировать Wnt7a, а вентральной поверхности — En1. Мишенями этих факторов являются клетки мезенхимы в зачатке конечности.
Передне-задняя ось. Зона поляризующей активности, расположенная на заднем крае зачатка конечности, участвует в организации пальцев. Важную роль в её спецификации играет гомеозисный ген Hox B8. Зона служит источником молекулярного сигнала Shh, который контролирует спецификацию передне-задней оси наряду с другими факторами, такими как Shh, FGF4, FGF8, BMP2 и BMP7.
Спецификация положения передних и задних конечностей находится под контролем транскрипционных факторов Tbx (T-box). В верхней конечности экспрессируется Tbx4, а в нижней — Tbx5. Продукты генов Tbx2 и Tbx3 найдены как в верхних, так и в нижних конечностях.
На поздних этапах развития конечности происходит ремоделирование хряща, формирование кости, образование скелетных мышц и их иннервация. Эти морфогенетические процессы связаны с массовой гибелью клеток, которая реализуется путём апоптоза. Хондрогенез, а также апоптоз контролируют BMP2 и BMP7. Их естественные антагонисты, факторы Gremlin и Noggin, сдерживают вступление клеток в апоптоз.
На 8-й неделе развития наблюдается ротация конечностей. Верхние и нижние конечности поворачиваются в различных направлениях: осуществляется супинация верхних конечностей и пронация нижних конечностей.
Нарушения развития конечностей
Мутации любого гена, контролирующего спецификацию структур в зачатке конечностей, приводит к развитию их многочисленных аномалий. В качестве примера приведём синдактилию (тип 2), связанную с нарушением экспрессии гомеозисного гена Hox 13.
Фармакологический препарат талидомид вызывал нарушения образования конечностей и был запрещён для использования с 1961 года. Тератогенное действие этого препарата связывают с его вмешательством в сигнальный механизм, опосредованный FGF8.
На ранних стадиях развития закладываются провизорные органы: хорион, амнион, желточный мешок и аллантоис. Они образуют оболочки зародыша, связывают его с организмом матери и выполняют некоторые специальные функции.
Бластоциста состоит из внутренней клеточной массы (эмбриобласт) и трофобласта (рис. 3-9, рис. 3-21). На 8–9-е сутки внутренняя клеточная масса расслаивается на эпибласт (первичная эктодерма) и гипобласт (первичная энтодерма). Kлетки гипобласта не принимают участия в образовании структур плода, их потомки присутствуют исключительно в составе провизорных органов (см. рис. 3-22, 3-23 и 3-24). Внезародышевая энтодерма формирует внутренний слой желточного мешка и аллантоиса. Внезародышевая эктодерма участвует в образовании внутреннего слоя амниона. Внезародышевая мезодерма разделяется на внутренний и наружный листки. Внутренний листок вместе с трофобластом формируют хорион, при этом клетки внезародышевой мезодермы обрастают трофобласт, образуя эндоцеломическую полость, или полость хориона. Наружный листок внезародышевой мезодермы участвует в формировании наружных слоёв амниона, желточного мешка и аллантоиса.
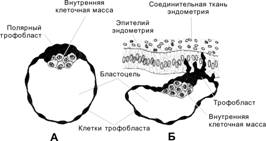
Рис. 3-21. Имплантация. А — бластоциста перед имплантацией (4,5 сут), Б — начало имплантации. Клетки трофобласта, расположенные на эмбриональном полюсе бластоцисты (полярная область), на 5–6 сут проникают в слизистую оболочку матки. [17]
Трофобласт
В трофобласте различают полярную область, покрывающую внутреннюю клеточную массу, и пристеночную (муральную) часть, образующую бластоцель. Клетки мурального трофобласта устанавливают контакт с материнской тканью в имплантационной крипте эндометрия матки и к моменту имплантации достигают состояния терминальной дифференцировки. В трофобласте развиваются два слоя: внутренний (цитотрофобласт) и наружный (синцитиотрофобласт).
Цитотрофобласт (слой Лангханса) состоит из интенсивно размножающихся клеток. Их ядра содержат хорошо различимые ядрышки, а клетки — многочисленные митохондрии, хорошо развитые гранулярную эндоплазматическую сеть и комплекс Гольджи. В цитоплазме содержится масса свободных рибосом и гранул гликогена.
Синцитиотрофобласт — высокоплоидная многоядерная структура, образуется из клеток цитотрофобласта. Служит источником плацентарного соматомаммотропина (плацентарный лактоген), хорионического гонадотропина (ХГТ) и эстрогенов. Митозы отсутствуют. Цитоплазма заполнена пузырьками различных размеров. Цитовиллин — периферический мембранный белок в синцитиотрофобласте.
В процессе формирования синцитиотрофобласта при слиянии клеток трофобласта происходит «фосфатидилсериновый переворот» (PS flip). Фосфатидилсерин (PS) — фосфолипид, присутствующий во внутреннем слое плазмолеммы. Перед слиянием клеток он перемещается в наружный слой плазмолеммы (фосфатидилсериновый переворот) и облегчает межмембранное слияние. Этот переворот является следствием активации каспаз (например, каспазы 8). В слиянии клеток трофобласта также участвует тканево-специфичный белок распознавания синцитин (гликопротеин, кодируемый геном HERV-W). В целом при слиянии клеток и формировании синцития происходит совместное действие синцитина и фосфолипидов наружного слоя плазматических мембран с участием вторичных посредников, таких как ионы Ca2+, инозитолтрифосфат и цАМФ.
Синцитин — гликопротеин слияния, кодируемый геном оболочки ретровируса, встроенным в геном человека (Env‑W ретровирусом). Синцитин, экспрессируемый в клетках трофобласта, индуцирует дифференцировку и слияние его клеток и формирование синцития (синцитиотрофобласта). Для экспрессии синцитина важны факторы транскрипции SP1 и GATA, которые индуцируют активность промотора синцитина. Дефекты синцитина могут обусловливать нарушения организации плаценты, приводящие к её дисфункции (например, преэклампсии).
В ходе слияния клеток и формировании синцития происходит формирование межклеточных щелевых контактов с участием преимущественно коннексина Cx43. Щелевые контакты обеспечивают диффузию молекул, регулирующих процесс слияния, между агрегированными клетками цитотрофобласта, а также между клетками цитотрофобласта и синцитиотрофобласта. В качестве регуляторных молекул могут выступать цАМФ, цГМФ, инозитолтрифосфат, ионы Ca2+. Экспрессия Cx43 и формирование щелевых контактов могут быть усилены аналогами цАМФ, глюкокортикоидами, хорионическим гонадотропином; как следствие, формирование ворсинок трофобласта происходит быстрее. С другой стороны, этот процесс угнетается эндотелином или трансформирующим фактором роста b.
Амнион
Амнион (рис. 3-22, 3-23) — плодный пузырь — объёмистый мешок, заполненный амниотической жидкостью (околоплодными водами). В области пупочного кольца амнион переходит на пуповину и далее на плодную часть плаценты, образуя их эпителиальный покров. Зародышевый (эмбриональный) и плодный периоды развития человека происходят внутри плодного пузыря.
Образование амниона
Одновременно с расслоением внутренней клеточной массы на эпибласт и гипобласт образуется амниотическая полость, ограниченная эпибластом и внезародышевой (амниотической) эктодермой. В ходе гаструляции клетки внезародышевой мезодермы обрастают амниотическую эктодерму, формируя наружный слой амниона.
Амниотические складки. На краниальном конце амнион образует головную амниотическую складку. С увеличением размеров зародыша его голова растёт вперёд в амниотическую складку, вогнутый край которой смещается в каудальном направлении. Боковые амниотические складки формируются по обе стороны зародыша за счёт краёв головной складки. Хвостовая амниотическая складка образуется на каудальном конце зародыша и растёт в краниальном направлении. Головная, боковые и хвостовая амниотические складки сходятся над зародышем и замыкают амниотическую полость. Место соединения амниотических складок — амниотический шов; здесь образуется исчезающий впоследствии тканевый тяж.
Амниотическая жидкость
Сформированный амниотический мешок наполняется жидкостью, защищающей зародыш при сотрясении, позволяющей плоду совершать движения и предотвращающей слипание растущих частей тела друг с другом и с окружающими тканями. Амниотическая жидкость играет важную роль в развитии пищеварительной, дыхательной и мочевыделительной систем. На 99% амниотическая жидкость состоит из воды, 1% приходится на белки, жиры, углеводы, ферменты, гормоны, неорганические соли. В амниотической жидкости обнаружены эпителиальные клетки амниона, кожи, кишечника, дыхательных и мочевыделительных путей, фибробласты, клетки мезенхимы. К концу беременности объём жидкости составляет 700–1000 мл.
Многоводие (hydramnios). Увеличение количества амниотической жидкости (2000 мл и более) возникает при нарушении заглатывания амниотической жидкости плодом. Многоводие сочетается с анэнцефалией и атрезией пищевода.
Маловодие (oligohydramnios). Уменьшение количества амниотической жидкости (менее 400 мл) часто связано с плацентарной недостаточностью. В случае агенезии почек маловодие обусловлено отсутствием вклада мочи в общий объём амниотической жидкости.
Образование и циркуляция амниотической жидкости
Хориоамниотическая мембрана. В результате увеличения плодного пузыря происходит постепенная облитерация внезародышевого целома (полость хориона). При этом амниотическая мембрана сливается с хорионом, образуя хориоамниотическую мембрану. Путём диффузии через хориоамниотическую мембрану из децидуальной оболочки в плодный пузырь поступает основной объём амниотической жидкости. Водные каналы, представленные аквапоринами 1, 8 и 9, обеспечивают реабсорбцию воды, поддерживая гомеостаз амниотической жидкости.
Почки. В амниотическую жидкость плод выделяет мочу, до 0,5 литра в день в последнем триместре беременности.
Лёгкие. Эпителий респираторного тракта в сутки выделяет 300–400 мл жидкости, поступающей в амнион. В плодном периоде лёгкие заполнены амниотической жидкостью, которая быстро резорбируется у новорождённого.
Кишечник. Плод заглатывает амниотическую жидкость (до 400 мл в сутки), которая таким образом попадает в кишечник, откуда происходит её абсорбция в кровь. В результате продукты обмена, поступающие в амниотическую жидкость из почек и лёгких, через плацентарный барьер выводятся в кровь матери.
Желточный мешок
Желточный мешок — вынесенная за пределы зародыша часть первичной кишки. Стенка желточного мешка состоит из двух слоёв: внутренний слой образован внезародышевой энтодермой, а наружный — внезародышевой мезодермой. В период наибольшего развития желточного мешка его кровеносные сосуды отделены от стенки матки тонким слоем ткани, что делает возможным поглощение из матки питательных веществ и кислорода. Внезародышевая мезодерма служит местом эмбрионального кроветворения (гемопоэза). Здесь формируются кровяные островки, в которых из гемангиобластов дифференцируются клетки крови и эндотелиальные клетки сосудов. Внезародышевая энтодерма желточного мешка служит временным местом локализации примордиальных половых клеток на пути их миграции в зачатки гонад, где они дифференцируются в гаметы. Позднее складки амниона сдавливают желточный мешок; образуется узкая перемычка, соединяющая его с полостью первичной кишки, — желточный стебелёк. Эта структура удлиняется и вступает в контакт с ножкой тела, содержащей аллантоис (рис. 3-23). Желточный стебелёк и дистальная часть аллантоиса вместе со своими сосудами образуют пупочный канатик, отходящий от зародыша в области пупочного кольца. Желточный стебелёк обычно полностью зарастает к концу 3-го месяца развития плода.
Дивертикул Меккеля. Приблизительно в 2% случаев (чаще у мужчин) часть желточного стебелька, прилегающая к кишке, образует дивертикул Меккеля — слепой отросток подвздошной кишки.
Дифференцировка гемангиобластов
Гемангиобласты, клетки предшественники стволовой кроветворной клетки и эндотелиальных клеток сосудов, дифференцируются из внезародышевой мезодермы. Морфоген Ihh, VEGF, ретиноевая кислота секретируются клетками внезародышевой энтодермы и стимулируют дифференцировку гемангиобластов, инициируя в клетках экспрессию факторов транскрипции (GATA1, GATA2, Tal-1), факторов гемопоэза (Lmo2), рецепторов VEGF (Flk1 и Flt1), антигенов дифференцировки (CD34, CD31).
VEGF
В формирующихся кровяных островках взаимодействие VEGF через рецептор Flk1 активирует в клетках-мишенях анти-апоптозный сигнальный каскад.
Ретиноевая кислота
Материнский витамин А (ретинол) в комплексе с ретинол-связывающим белком путём опосредованного рецептором эндоцитоза поступает в клетки внезародышевой энтодермы. Внутриклеточный ретинол-связывающий белок CRBPI и ретинол дегидрогеназы (RoDH и Raldh2) обеспечивают синтез ретиноевой кислоты. В эндотелиальных клетках мезодермы ретиноевая кислота через рецепторы a1 и a2 стимулирует экспрессию фактора роста TGFb-1, молекулы межклеточного матрикса фибронектина, молекулы адгезии a5 интегрина. Хромосомная аномалия, затрагивающая ген рецептора ретиноидов, может стать причиной острой промиелоцитозной лейкемии. Транслокация затрагивает 15 и 17 хромосомы, t(15;17). Эпидемиологические наблюдения обнаружили дефект гемопоэза при недостаточном поступлении в организм витамина А.
Дифференцировка примордиальных половых клеток
На второй неделе эмбрионального развития в проксимальной части эпибласта дифференцируется примерно 40 примордиальных половых клеток. Морфогены BMP4 и BMP8b, секретируемые клетками внезародышевой эктодермы, и BMP2, продуцируемый клетками внезародышевой энтодермы, контролируют дифференцировку примордиальных половых клеток. Репрессор транскрипции Blimp1 регулирует дифференциальную активность гомеозисных генов, экспрессируемых в половых клетках. В ходе гаструляции экспрессия Ifitm/mil/fragilis (индуцированный интерфероном трансмембранный белок) обеспечивает миграцию клеток из эпибласта через первичную полоску в энтодерму желточного мешка по принципу репульсивного (отталкивающего) механизма. В ходе миграции половые клетки «сторонятся» клеток мезодермы, также экспрессирующих Ifitm/mil/fragilis. На 4-й неделе развития примордиальные половые клетки из желточного мешка по ходу брыжейки мигрируют в направлении половых валиков.
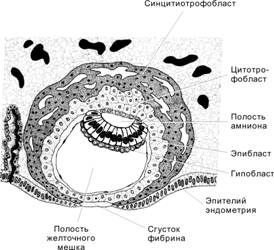
Рис. 3-22. Погружённая в эндометрий бластоциста. Внутренняя часть трофобласта представлена цитотрофобластом. В наружной части трофобласта (синцитиотрофобласт) имеется большое количество лакун. Внутренняя клеточная масса расслоилась на эпи- и гипобласт. [17]
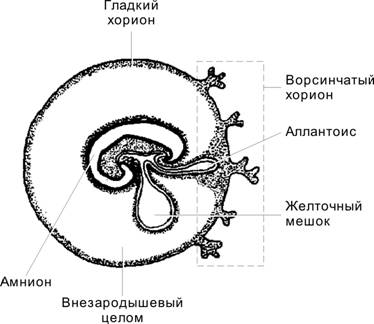
Рис. 3-23. Внезародышевые оболочки. Зародыш находится в полости амниона. Желточный мешок связан с зачатком пищеварительной системы, а с каудальной её частью сообщается аллантоис. Все эти структуры находятся в эндоцеломической полости. Хорион топографически и структурно разделяется на гладкий и ворсинчатый. [17]
Аллантоис
Задняя стенка желточного мешка к 16-му дню развития формирует небольшой вырост — аллантоис (греч. аllas, колбасовидный), образованный внезародышевыми энтодермой и мезодермой. Устье аллантоиса открывается в клоаку, а его дистальная часть проникает в ножку тела (рис. 3-23). У человека аллантоис рудиментарен, он не функционирует как орган дыхания или резервуар для окончательных продуктов обмена у животных, но имеет важное значение в эмбриональном кроветворении и ангиогенезе. На 3–5-й неделе развития в стенке аллантоиса происходит гемопоэз и формируются кровеносные сосуды пупочного канатика (две пупочные артерии и одна пупочная вена). Клетки внезародышевой мезодермы экспрессируют специфические для гемопоэза факторы транскрипции (GATA1, GATA2, Tal-1), рецепторы VEGF. На 7-й неделе эмбриогенеза уроректальная перегородка разделяет клоаку на прямую кишку и мочеполовой синус, соединённый с аллантоисом (см. рис. 12-42). Поэтому проксимальный отдел аллантоиса имеет отношение к образованию мочевого пузыря, что следует учитывать при аномалиях развития этого органа. На 2-м месяце эмбриогенеза аллантоис дегенерирует, а на его месте появляется урахус ¾ плотный фиброзный тяж, тянущийся от верхушки мочевого пузыря к пупочному кольцу. В постнатальном периоде урахус организуется в срединную пупочную связку.
Аномалии урахуса
Пупочно-пузырные свищи. При полном незаращении просвета урахуса сохраняется его связь с полостью мочевого пузыря, что является причиной выделения мочи из пупка.
Киста урахуса. Частичная облитерация урахуса приводит к образованию замкнутых полостей, заполненных инфицированной мочой.
Опухоли урахуса встречаются крайне редко.
Плацента
Плацента связывает плод с организмом матери. Плацента состоит из материнской (базальная часть децидуальной оболочки) и плодной (ворсинчатый хорион — производное трофобласта и внезародышевой мезодермы) частей (рис. 3-24А). В полости матки бластоциста находится в течение 1,5–2 суток. Имплантация происходит на 20–21-й день нормального менструального цикла, или через 5,5–6 суток после оплодотворения, когда функциональный слой эндометрия имеет наибольшую толщину (рис. 3-24). В ходе имплантации бластоциста вступает в тесный контакт с эпителием слизистой оболочки матки. В некоторых местах соприкасающиеся эпителиальные клетки матки и клетки трофобласта формируют специализированные контакты, в т.ч. щелевые. Под действием маточного секрета прозрачная оболочка растворяется. После этого бластоциста прикрепляется к эндометрию, как правило, тем полюсом, на котором расположена внутренняя клеточная масса (полярный трофобласт). Через двое суток бластоциста оказывается полностью погружённой в слизистую оболочку матки.
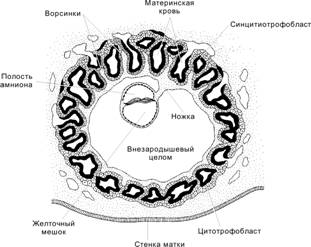
Рис. 3-24. Трёхнедельный эмбрион на стадии гаструлы. Сформированы полость амниона и желточный мешок. Клетки трофобласта, образующие плаценту, вступают в контакт с кровеносными сосудами матки. Зародыш связан с трофобластом происходящей из внезародышевой мезодермы ножкой тела. В ножку тела прорастает аллантоис, здесь протекает ангиогенез, и в дальнейшем формируется пупочный канатик с проходящими в его составе пупочными (аллантоисными) сосудами: двумя пупочными артериями и одной пупочной веной. [17]
Рис. 3-24А. Плацента. Ворсинки хориона (1) снаружи покрывает многоядерная структура — синцитиотрофобласт (4). Межворсинчатое пространство (2) заполнено материнской кровью. Сердцевина ворсинки образована соединительной тканью, содержащей кровеносные сосуды (3). Окраска гематоксилином и эозином.
Хорион
Соединение трофобласта и внезародышевой мезодермы приводит к образованию хориона. В формировании хориона различают три периода: предворсинчатый (7–8-й день развития), период образования ворсинок (до 50-го дня), период котиледонов (с 50-го по 90-й день).
Предворсинчатый период
В ходе имплантации клетки трофобласта пролиферируют и образуют цитотрофобласт (рис. 3-24, 3-25), снаружи от которого расположен синцитий — производное цитотрофобласта. На ранних сроках имплантации трофобласт не обладает выраженными цитолитическими свойствами: бластоциста проникает между клетками поверхностного эпителия без его разрушения. В дальнейшем, по мере взаимодействия с эндометрием, трофобласт проявляет цитолитическую активность. В ткани эндометрия, окружающей трофобласт, появляются полости (лакуны), заполненные кровью матери. Лакуны разделены перегородками из клеток трофобласта (первичные ворсинки). После появления лакун бластоциста может быть названа плодным пузырём; наступает ворсинчатый период развития плаценты.

Рис. 3-25. Развитие ворсинок хориона. А — первичная, Б — вторичная, В — третичная ворсинка. [17]
Период образования ворсинок
В этот период последовательно образуются первичные, вторичные и третичные ворсинки (рис. 3-25).
Первичные ворсинки — скопления клеток цитотрофобласта, окружённые синцитиотрофобластом. Клетки цитотрофобласта имеют неправильную форму и светлую цитоплазму.
Вторичные ворсинки. На 12–13-й день в первичные ворсинки врастает внезародышевая мезодерма, что приводит к формированию вторичных ворсинок, равномерно распределённых по всей поверхности плодного яйца. Вторичные ворсинки имеют наибольшую длину в базальной части децидуальной оболочки. Эпителий вторичных ворсинок представлен светлыми клетками округлой формы с крупными ядрами. Над эпителием расположен синцитий с нечёткими границами, тёмной зернистой цитоплазмой, щёточной каёмкой и полиморфными ядрами. На стадии вторичных ворсинок в клетках трофобласта уменьшается число митозов, и начинается их васкуляризация.
Третичные ворсинки. С 3-й недели развития появляются третичные ворсинки, содержащие кровеносные сосуды. Этот период называют плацентацией. Ворсинки, обращённые к базальной части децидуальной оболочки, кровоснабжаются не только из сосудов, происходящих из хориальной мезодермы, но и из сосудов аллантоиса. Период соединения ветвей пупочных сосудов с местной сетью кровообращения совпадает с началом сердечных сокращений (21-й день развития), и в третичных ворсинках начинается циркуляция эмбриональной крови. Васкуляризация ворсинок хориона заканчивается, как правило, на 10-й неделе беременности. К этому сроку формируется плацентарный барьер. Не все ворсинки хориона развиты одинаково хорошо. Ворсинки, обращённые к капсулярной части отпадающей оболочки, развиты слабо и постепенно исчезают. Поэтому хорион в этой части называют гладким. Строма гладкого хориона, обращённого к капсулярной части децидуальной оболочки, бедна кровеносными сосудами.
Период котиледонов
Котиледон — структурно-функциональная единица сформированной плаценты. Он образован стволовой ворсинкой и её разветвлениями, содержащими сосуды плода. К 140-му дню беременности в плаценте сформировано 10–12 больших, 40–50 мелких и до 150 рудиментарных котиледонов. К 4-му месяцу беременности формирование основных структур плаценты заканчивается (рис. 3-26). Лакуны полностью сформированной плаценты содержат около 150 мл материнской крови, полностью замещаемой 3–4 раза в минуту. Oбщая поверхность ворсинок достигает 14 м2, что обеспечивает высокий уровень обмена между беременной и плодом.
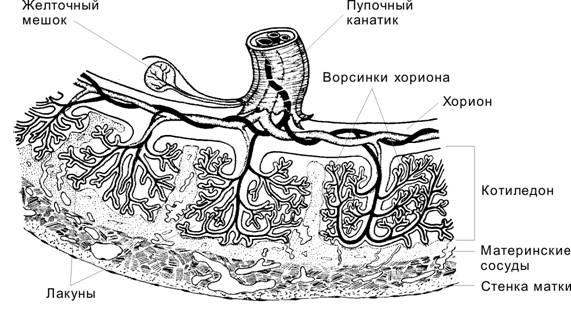
Рис. 3-26. Плацента. С увеличением срока беременности ворсинки становятся разветвлённее. В сформированной плаценте присутствуют скопления ворсинок — котиледоны. Часть ворсинок (закрепляющие, или якорные) более тесно связана с материнской тканью. [17]
Децидуальная оболочка
Децидуальная (отпадающая) оболочка образуется на всём протяжении эндометрия, но раньше она развивается в области имплантации. К концу второй недели эндометрий полностью замещается децидуальной оболочкой, в которой различают базальную (decidua basalis), капсулярную (decidua capsularis) и пристеночную (decidua parietalis) части. Децидуальная оболочка, окружающая хорион, образует базальную и капсулярную части (рис. 3-27). В остальных областях полость матки выстлана пристеночной децидуальной оболочкой. В децидуальной оболочке выделяют губчатую (спонгиозную) и компактную зоны. Более рыхлая губчатая зона содержит остатки желёз эндометрия.

Рис. 3-27. Децидуальная оболочка матки и плацента. Полость матки выстилает пристеночная часть децидуальной оболочки. Капсулярная и базальная её части покрывают хорион. Базальная часть децидуальной оболочки, обращённая к ворсинчатому хориону, входит в состав плаценты. [17]
Базальная часть децидуальной оболочки входит в состав плаценты (рис. 3-27). Она отделяет плодное яйцо от миометрия. В губчатом слое много желёз, сохраняющихся до 6-го месяца беременности.
Капсулярная часть. К 18-му дню беременности капсулярная часть полностью смыкается над имплантированным плодным яйцом и отделяет его от полости матки. По мере развития плода капсулярная часть выпячивается в полость матки и к 16-й неделе срастается с пристеночной частью. При доношенной беременности капсулярная часть хорошо различима лишь в нижнем полюсе плодного яйца (над внутренним зевом). Поверхностного эпителия в капсулярной части нет.
Пристеночная часть. До 15-й недели беременности пристеночная часть децидуальной оболочки утолщается за счёт компактной и губчатой зон. Позже в компактной зоне исчезают крупные децидуальные клетки. В губчатой зоне пристеночной части децидуальной оболочки железы развиваются до 8-й недели беременности. K моменту слияния пристеночной и капсулярной частей количество желёз постепенно уменьшается, они становятся неразличимыми. В конце доношенной беременности пристеночная часть децидуальной оболочки представлена несколькими слоями децидуальных клеток. С 12-й недели беременности поверхностный эпителий пристеночной части исчезает.
Децидуальные клетки. Клетки рыхлой соединительной ткани вокруг сосудов компактной зоны резко увеличены. Это молодые децидуальные клетки, сходные с фибробластами. По мере дифференцировки размеры децидуальных клеток увеличиваются, они приобретают округлую форму, их ядра становятся светлыми, клетки теснее прилегают друг к другу. К 4–6-й неделе беременности преобладают крупные децидуальные клетки. Часть децидуальных клеток имеет костномозговое происхождение; по-видимому, они участвуют в иммунном ответе. Децидуальные клетки продуцируют пролактин и простагландины.
Ворсинчатый хорион
Третичные ворсинки хорошо развиты на стороне, обращённой к миометрию, где образуется ворсинчатый хорион. В ворсинчатом хорионе капилляры приближены к базальной мембране трофобласта и образуют поверхностную капиллярную сеть. Среди клеточных элементов стромы третичных ворсинок присутствуют макрофаги — клетки Хофбауэра. От вершины ворсинок в сторону децидуальной ткани отходят клеточные столбики (колонны), состоящие из клеток цитотрофобласта, контактирующих с поверхностной компактной зоной децидуальной оболочки. В области контакта формируется зона коагуляционного некроза (слой Нитабух). Далее клетки цитотрофобласта проникают в губчатую зону эндометрия, миометрий и стенку сосудов матки. Врастание цитотрофобласта на 6-й неделе беременности в стенку спиральных артерий приводит к вскрытию их просвета и установлению циркуляции материнской крови между ворсинками хориона. Ворсинки, тесно связанные с материнской тканью, называют якорными, или закрепляющими. Но большинство ворсинок в полостях базальной части децидуальной оболочки лежит более или менее свободно.
Молекулярные механизмы имплантации и развития плаценты
В предимплантационный период клетки эндометрия экспрессируют A-рецепторы прогестерона и a-рецепторы эстрогена. Эстроген, связываясь со своими рецепторами, инициирует формирование рецептивного поля для бластоцисты в эндометрии, а также стимулирует образование в эндометрии ряда ростовых факторов, наиболее значимым из которых является лейкоз ингибирующий фактор (LIF, Leukemia Inhibitory Factor). LIF играет важную роль в запуске и нормальном ходе имплантации. LIF продуцируется клетками эпителия и желёз эндометрия в середине секреторной фазы менструального цикла, а во время имплантации — клетками стромы эндометрия.
Первый этап в ходе имплантации — слабое и нестабильное связывание бластоцисты с эпителием рецептивного участка примированного гормонами эндометрия. При этом бластоциста освобождается от прозрачной оболочки, а клетки трофобласта начинают экспрессировать L-селектин. Одновременно в эпителиальных клетках эндометрия рецептивного участка активируются олигосахаридные лиганды, они связываются с L-селектином клеток трофобласта. Такое связывание относительно нестабильно. Затем слабое связывание становится прочным, что позволяет трофобласту внедряться в ткань эндометрия.
В прочном связывании с эндометрием участвуют интегрины. Так, экспрессируемый трофобластом anb3 интегрин связывается с фибронектином и перлеканом (гепаран сульфат протеогликаном) внеклеточного матрикса эндометрия. anb3 Интегрин экспрессируется также клетками эпителия эндометрия в 19–24 сутки менструального цикла, когда рецептивность эндометрия для бластоцисты оптимальна. У женщин с дефектами лютеиновой фазы овариально-менструального цикла отмечено отсутствие anb3 интегрина в эпителии эндометрия, что ведёт к бесплодию.
LIF, фактор роста эпидермиса, интерлейкин-1, инсулиноподобные факторы роста, в присутствии Hox A10 регулируют активность циклооксигеназы и образование простагландинов, необходимых для имплантации. Циклооксигеназа (COX) существует в двух изоформах: конститутивной (COX-1) и индуцибельной (COX-2). В эндометрии прогестерон и эстроген подавляют образование COX-1 в середине лютеиновой фазы овариального цикла, тогда как образование COX-2 сохраняется. При дефектах COX-2 и отсутствии простагландинов (особенно простациклина) имплантации не происходит. Простагландин I2, продуцируемый с участием COX-2, является лигандом для PPAR. Ядерный рецептор PPARg (Peroxisome Proliferator-Activated Receptor g) важен для имплантации, дифференцировки и функционирования трофобласта; дефекты PPARg ведут к аномальному развитию плаценты.
В ходе имплантации продукция цитотрофобластом протеиназ (в частности, металлопротеиназы-9) и их активность нарастают; усилению образования и активности металлопротеиназы-9 способствует ИЛ1, продуцируемый эндометрием.
В ходе формирования ворсин плаценты клетки цитотрофобласта экспрессируют фактор glial cells missing-1 (контролирующий переход нейрон–глия у Drosophila).
В развитии децидуальной реакции участвуют интерлейкин-11, инсулиноподобные факторы роста, Hox A10, фактор транскрипции forkhead.
В дифференцировке цитотрофобласта и инвазии в эндометрий большую роль играет содержание кислорода. Кислород контролирует баланс между пролиферацией цитотрофобласта и его дифференцировкой. Ранние события развития плаценты происходят в условиях относительной гипоксии (2–5% O2), что является благоприятным условием для пролиферации, но не дифференцировки цитотрофобласта в ходе имплантации; количество клеток в составе цитотрофобласта быстро увеличивается. Клетки цитотрофобласта внедряются глубже в стенку матки и перекрывают просвет материнских сосудов, поддерживая тем самым состояние физиологической гипоксии. Таким образом, в течение первого триместра беременности наблюдается преобладающий рост цитотрофобласта по сравнению с ростом плода, сохраняющего сравнительно небольшие размеры, то есть формирование плаценты предваряет рост плода. Во вторую половину беременности, при наличии зрелой функционирующей плаценты, происходит быстрый рост плода.
Реакции клеток на недостаток O2 контролируются индуцированным гипоксией фактором HIF (Hypoxia-Inducible Factor). Три белка семейства HIFa являются факторами транскрипции bHLH, которые содержат Per/Arnt/Sim домен, способствующий димеризации HIFa с HIFb. Гетеродимеры активируют транскрипцию ряда важных генов, в промоторе которых присутствует HIF-чувствительный элемент. По мере углубления в стенку матки и внедрения в материнские сосуды клетки трофобласта получают доступ к большему количеству кислорода, который опосредует их выход из клеточного цикла и последующую дифференцировку.
К 10–12 неделям беременности начинается ток крови в межворсинчатых пространствах. По мере эндоваскулярной инвазии цитотрофобласта клетки мигрируют вдоль спиральных артерий, замещая эндотелий, а также проникая в их мышечную оболочку. Просвет артерий значительно увеличивается. Это усиливает кровоснабжение, что необходимо для поддержания быстрого роста плода на более поздних сроках беременности. Недостаточность внутрисосудистой инвазии трофобласта может привести к аборту; неспособность к внедрению на необходимую глубину сдерживает роста плода.
Многие обычные препараты (например, никотин) отрицательно влияют на дифференцировку трофобласта и взаимоотношение между матерью и плодом, в том числе на плацентарный транспорт аминокислот и рост плода. Субклинические вирусные (например, цитомегаловирус) или бактериальные инфекции могут ингибировать дифференцировку цитотрофобласта и/или внедрение (инвазию) в эндометрий.
В случае трисомии 21 нарушена способность клеток цитотрофобласта сливаться и формировать синцитиотрофобласт. При этом снижается инвазивная активность трофобласта и отмечено увеличение количества его клеток, вступающих в апоптоз. Подобные дефекты могут быть причиной аборта. В других случаях, например при плацентарном мозаицизме, который выявлен в 2% беременностей, закончившихся живым плодом, отмечено частое ограничение роста плода.
Организация провизорных органов у близнецов имеет ряд особенностей. Eсли близнецы разнояйцовые (дизиготные, 70% всех близнецов), то в этом случае каждый зародыш имеет собственные амнион, хорион и плаценту; плаценты иногда сливаются (рис. 3-28). Oднояйцовые (монозиготные) близнецы обычно имеют собственные амнион и общие хорион и плаценту. B случае неразделённых или не полностью разделённых близнецов присутствует один амнион, один хорион и одна плацента (рис. 3-29).
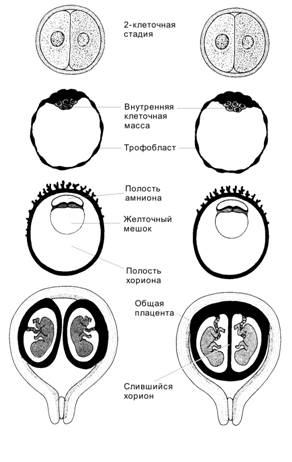
Рис. 3-28. Развитие разнояйцовых близнецов. [17]

Рис. 3-29. Развитие однояйцовых близнецов. [17]
Функция плаценты
Обмен между матерью и плодом газами, метаболитами, электролитами реализуют пассивный транспорт, облегчённая диффузия и активный транспорт. Через плаценту в организм плода свободно проходят стероидные гормоны.
Транспорт материнских АТ осуществляется при помощи опосредованного рецепторами эндоцитоза и обеспечивает пассивный иммунитет плода.
Эндокринная функция. Плацента — эндокринный орган. Плацента синтезирует ряд гормонов и других биологически активных веществ, имеющих важное значение для нормального течения беременности и развития плода (ХГТ прогестерон, хорионический соматомаммотропин, фактор роста фибробластов, трансферрин, пролактин и релаксин). Кортиколиберин, вероятно, определяет срок наступления родов.
Плацентарные часы. При изучении содержания в крови 485 беременных плацентарного кортиколиберина установлено, что уже на сроке 18 недель беременности можно выделить группы с нормальными, преждевременными и поздними родами. К предсказанному для этих групп сроку содержание в крови кортиколиберина резко возрастает.
Детоксикация некоторых лекарственных препаратов.
Плацентарный барьер. В состав плацентарного барьера (материнская кровь — кровь плода) входят: синцитиотрофобласт ® цитотрофобласт ® базальная мембрана трофобласта ® соединительная ткань ворсинки ® базальная мембрана в стенке капилляра плода ® эндотелий капилляра плода. Хорион защищает плод от влияния иммунной системы матери.
· Угнетение клеточного иммунного ответа. Согласно одной из гипотез, хорион вырабатывает вещества, угнетающие клеточный иммунный ответ. В пользу этого предположения свидетельствуют данные о том, что ферменты мембран, а также экстракт из синцитиотрофобласта тормозят in vitro размножение клеток иммунной системы матери.
· Плацентарный барьер и HLA. Другой механизм защиты плода от атакующего воздействия лимфоцитов матери связывают с отсутствием Аг HLA в трофобласте человека, что угнетает развитие цитотоксического иммунного ответа со стороны матери. Отсутствие Аг HLA в трофобласте обеспечивает защиту фетоплацентарного комплекса от распознавания иммунокомпетентными клетками матери. Другие типы клеток в ворсинках плаценты человека несут Аг HLA. Трофобласт человека, наряду с отсутствием Аг HLA, не содержит также эритроцитарных Аг систем AB0 и Rh.
Врождённые пороки (ВП), включая аномалии развития, их причины изучает тератология. Число новорождённых с ВП составляет 2–3% общего количества родившихся живыми детей. Причины ВП многочисленны: вирусная инфекция (краснуха, цитомегаловирусная и герпетическая инфекции), токсоплазмоз, сифилис, радиация, лекарственные препараты, наркотические вещества, химические факторы окружающей среды, болезни матери и т.д. Восприимчивость к действию тератогенов зависит от стадии развития. Риск возникновения ВП особенно велик в эмбриогенезе и органогенезе. Воздействие тератогенов в первые 3 недели развития заканчивается гибелью эмбриона с последующим спонтанным абортом. Критический период возникновения врождённых пороков приходится на 3–8-ю неделю развития, с пиком на 4–5-й неделе (табл. 3-4). Большинство ВП обусловлено воздействием факторов внешней среды, генетическими дефектами или их сочетанием. В ряде случаев не удаётся установить причину врождённого дефекта (спорадические заболевания).
Таблица 3-4. Критические периоды возникновения врождённых пороков и частота выявляемых аномалий на 1000 живорождённых
|
Орган |
Критический период (недели развития) |
Частота аномалий (на 1000 плодов) |
|
ЦНС |
3–16 |
10 |
|
Сердце |
3–6 |
8 |
|
Почки |
5–8 |
4 |
|
Конечности |
4–5 |
2 |
|
Верхняя губа |
5–6 |
6 |
|
Нёбо |
7–8 |
|
|
Половые органы |
7–9 |
|
|
Всего |
30 |
|
Дисплазии
Дисплазии — морфологические изменения, выходящие за пределы общепринятой нормы; подразделяют на мальформации, деформации и дизрупции.
Мальформации — ВП, возникающие при неправильном формировании структур (проявления генных или хромосомных аномалий, многофакторные заболевания, эффект тератогенов).
Деформации — ВП, возникающие в результате механического воздействия на нормально формирующийся плод. К деформациям могут привести аномалии строения матки, маловодие, многоплодная беременность, фиброз матки. К развитию деформаций может привести также недостаточная подвижность плода (например, нервно-мышечные заболевания, аномалии внутриутробного расположения плода).
Дизрупции (разрушения) — ВП, возникающие в нормально развивающихся органах под воздействием инфекционных агентов, механических повреждений (амниотические перетяжки) или сосудистой окклюзии.
Причины
Более половины ВП (50–60%) имеют неизвестную этиологию. Среди установленных причин возникновения ВП 7–10% обусловлены воздействием факторов окружающей среды, 7–8% — генные мутации, 6–7% — хромосомные аберрации и 20–25% ВП вызваны сочетанными причинами.
Тератогенные воздействия
По крайней мере, 10% всех ВП обусловлено воздействием факторов внешней среды. Факторы окружающей среды, вызывающие ВП, получили название тератогенов. Вещества, загрязняющие окружающую среду, можно рассматривать как тератогены. Эффект тератогенов обусловлен влиянием на гисто- и органогенез, рост и развитие плода. Доза тератогенов имеет существенное значение. Для многих тератогенов существует порог концентрации, ниже которого статистическая вероятность тератогенного эффекта ничтожно мала. Генетическая конституция матери и особенно плода во многом определяет, окажется ли данное воздействие тератогенным. Например, только в 11% случаев после приёма дифенилгидантоина во время беременности развивается гидантоиновый синдром плода.
Инфекционные агенты попадают в кровь плода через плацентарный барьер. Инфицирование плода может закончиться абортом или возникновением ВП (табл. 3-5).
Таблица 3-5. Инфекционные агенты, обладающие тератогенными эффектами
|
Возбудитель |
Врождённые пороки |
|
Вирус краснухи (возбудитель кори) |
Катаракта, глухота, задержка умственного развития, ВП сердца |
|
Цитомегаловирус |
Нарушение роста плода, микроцефалия, задержка умственного развития, глухота, слепота, гепатоспленомегалия |
|
Вирус герпеса |
Микроцефалия, микрофтальмия, дисплазия сетчатки, задержка умственного развития |
|
Бледная трепонема (возбудитель сифилиса) |
Умственная отсталость, менингиты, гидроцефалия, глухота, дефекты развития зубов и лица |
|
Простейшее Toxoplasma gondii |
Микроцефалия, микрофтальмия, гидроцефалия, задержка умственного развития |
Высокие дозы радиации — тератогенный фактор, способный вызывать ВП (микроцефалия, задержка умственного развития, аномалии скелета, катаракта), нарушая деление клеток и органогенез.
Медицинские препараты и химические вещества. Антиконвульсанты, антикоагулянты, антитиреоидные и психотропные препараты, антибластомные химиопрепараты, йодсодержащие вещества, гидантоин, талидомид, варфарин, диэтилстильбэстрол, противозачаточные препараты могут вызвать разные пороки развития (табл. 3-6).
Таблица 3-6. Врождённые пороки, обусловленные воздействием различных веществ
|
Тератоген |
Врождённые пороки |
|
Алкоголь |
Нарушает рост плода, приводит к развитию аномалий головного мозга, ВП сердца, скелета. Женщины, страдающие хроническим алкоголизмом, имеют угрозу рождения ребёнка с комплексом ВП (алкогольный синдром плода) |
|
Ретиноевая кислота (витамин А) |
Дисплазия лица, аномалии головного мозга и сердца |
|
Тетрациклин |
Гипоплазия эмали и образование тёмных пигментных пятен на поверхности зубов |
|
Стрептомицин |
Потеря слуха, повреждения слухового нерва |
|
Ртуть |
Аномалии головного мозга, задержка умственного развития, слепота, спастичность |
|
Литий |
Дефекты слуха |
Метаболические нарушения у матери
· Сахарный диабет. Дети, рождённые женщинами, страдающими сахарным диабетом, имеют риск возникновения ВП (особенно сердца, скелета, ЦНС) около 10–15%. Видимо, основной тератогенный фактор — гипергликемия.
· Фенилкетонурия. Дети, рождённые женщинами, страдающими фенилкетонурией, внутриутробно подвергались воздействию избыточной концентрации метаболитов фенилаланина; при этом почти всегда формируются ВП сердца и дефекты ЦНС.
Генетические нарушения
Генетические факторы могут приводить как к единичным ВП, так и к развитию многообразных синдромов. Хромосомные аберрации (мозаицизм, аномалии числа и структуры хромосом) чаще возникают в результате мутаций de novo. Дефекты генов, кодирующих конкретный белок, передаются по аутосомно-доминантному, аутосомно-рецессивному или сцепленным с полом типу наследования.
Спорадические заболевания
Они часто бывают следствием нарушения эмбрионального развития при патологическом течении беременности (например, при окклюзии кровеносных сосудов). Некоторые врождённые аномалии могут возникать в результате спонтанной доминантной мутации (Â) соматических клеток, приводящей либо к летальному исходу, либо оказывающей воздействие на репродуктивную функцию.
Пренатальная диагностика
Раннее выявление ВП или генетического заболевания у плода позволяет либо принять решение о прерывании беременности, либо морально подготовиться к рождению больного ребёнка.
Амниоцентез
Обычно амниоцентез проводят на 16-й неделе беременности. Путём прокола плодного пузыря получают 20–30 мл амниотической жидкости для цитогенетического и биохимического анализа.
Цитогенетическое исследование
· Хромосомный анализ. Для выявления аномалий хромосомы окрашивают красителем Гимзы.
Определение пола плода имеет важное диагностическое значение при угрозе наследования заболевания, сцепленного с полом. Если мать — носитель заболевания, сцепленного с Х-хромосомой, то прерывание беременности при установлении мужского пола плода позволит избежать рождения больного ребёнка).
· Прямое исследование ДНК проводится, если имеется риск возникновения моногенного заболевания с известным генным дефектом. Генное зондирование (например, ПЦР) позволяет обнаружить точечные мутации генов-мишеней.
Биохимические исследования
· a-Фетопротеин сыворотки крови матери; уровень определяют на 16–18 неделе беременности. Повышенное содержание a-фетопротеина указывает на возможные пороки развития ЦНС, дефекты передней брюшной стенки (например, омфалоцеле), отслойку плаценты. Значимым принято считать повышение содержания a-фетопротеина в 2,5 раза и более от среднего значения для конкретного срока беременности. Понижение уровня a-фетопротеина наблюдают при болезни Дауна и других трисомиях. Значимо уменьшение уровня a-фетопротеина вдвое против средних значений для конкретного срока беременности (особенно в сочетании с повышением содержания другого сывороточного маркёра — ХГТ).
· Сурфактант. Отношение содержания лецитина к содержанию сфингомиелина в амниотической жидкости косвенно характеризует количество внутриальвеолярного сурфактанта и степень зрелости лёгких. Показатель 2:1 или выше — признак функциональной зрелости лёгких.
Биопсия ворсин хориона
Пункцию ворсин хориона проводят на 8–11 неделе беременности. Полученные аспирацией клетки ворсин хориона подвергают цитогенетическому и биохимическому исследованию. Преимуществом пункции ворсин хориона, по сравнению с амниоцентезом, являются сроки выполнения исследований, биопсия ворсин хориона проводится на несколько недель раньше, чем амниоцентез.
Ультразвуковое исследование
УЗИ — безопасный метод пренатальной диагностики. Считают, что УЗИ не представляет какого-либо риска ни для матери, ни для плода. При УЗИ обычно определяют размеры и массу плода, их количество и жизнеспособность. Также оценивают состояние плаценты, пуповины, околоплодных вод. При УЗИ можно обнаружить большинство структурных аномалий основных органов и систем. В этих случаях необходимо проведение амниоцентеза с биопсией ворсин хориона.
Фетоскопия